Insulin
| Insulin | ||
|---|---|---|
| ||
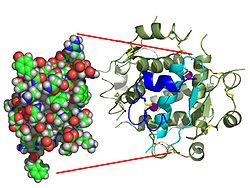
| Zwei Modelldarstellungen des Insulinmoleküls. Links das einfache Molekül (Monomer) als ein Kalottenmodell aus dem die Oberflächenform hervorgeht. Rechts das sechsfache Molekül (Hexamer) als so genanntes Bändermodell, bei dem die innere Struktur deutlich wird. Bei letzterem sind α-Helices durch Schrauben und β-Faltblätter durch Pfeile dargestellt. | ||
| Vorhandene Strukturdaten: 1ai0, 3ins, 4ins, 6ins, 7ins, 9ins | ||
| Eigenschaften des menschlichen Proteins | ||
| Masse/Länge Primärstruktur | 5,8 kDa / 51 Aminosäuren | |
| Sekundär- bis Quartärstruktur | Heterodimer (21 + 30 aa) | |
| Präkursor | Proinsulin | |
| Bezeichner | ||
| Gen-Name | ||
| Externe IDs |
| |
| Arzneistoffangaben | ||
| ATC-Code | A10AB01 | |
| DrugBank | DB00030 | |
| Vorkommen | ||
| Homologie-Familie | HBG006137 | |
| Übergeordnetes Taxon | Wirbeltiere | |
| Orthologe | ||
| Mensch | Maus | |
| Entrez | 3630 | 16334 |
| Ensembl | ENSG00000129965 | ENSMUSG00000000215 |
| UniProt | P01308 | Q5EEX1 |
| Refseq (mRNA) | NM_000207 | NM_008387 |
| Refseq (Protein) | NP_000198 | NP_032413 |
| Genlocus | Chr 11: 2.14 – 2.14 Mb | Chr 7: 142.49 – 142.49 Mb |
| PubMed-Suche | 3630 | 16334 |

Insulin (andere Namen: Insulinum, Insulinhormon, Inselhormon) ist ein für alle Wirbeltiere lebenswichtiges Proteohormon (Polypeptidhormon), das in den β-Zellen der Bauchspeicheldrüse gebildet wird. Diese spezialisierten Zellen befinden sich nur in den Langerhans-Inseln. Von diesen Inseln leitet sich auch der Name „Insulin“ ab (lateinisch insula). Insulin senkt den Blutzuckerspiegel, indem es Körperzellen dazu anregt, Glukose aus dem Blut aufzunehmen.
Funktion und Wirkung
Die Regulation der Konzentration von Glukose im Blut erfolgt durch einen Regelkreis aus zwei Hormonen, die abhängig von der Blutzuckerkonzentration ausgeschüttet werden. Insulin ist das einzige Hormon, das den Blutzuckerspiegel senken kann. Sein Gegenspieler ist das Glucagon, dessen Hauptaufgabe es ist, den Blutzuckerspiegel zu erhöhen. Auch Adrenalin, Kortisol und Schilddrüsenhormone haben blutzuckersteigernde Wirkungen.
Der Blutzuckerspiegel steigt vor allem nach der Aufnahme von kohlenhydratreicher Nahrung. Als Reaktion darauf wird von den β-Zellen Insulin ins Blut ausgeschüttet. Insulin senkt den Blutzuckerspiegel dadurch, dass es mittels seiner „Schlüsselfunktion“ der Glukose aus dem Blutplasma und der Gewebsflüssigkeit den Durchtritt durch die Zellmembran in das Zellinnere ermöglicht. Vor allem die Leber- und Muskelzellen können in kurzer Zeit große Mengen Glukose aufnehmen und sie in Form von Glykogen speichern oder in Energie umwandeln (siehe Glykolyse).
Auch auf andere Zellen wirkt das Hormon, so hat es Einfluss auf den Fett- und Aminosäurestoffwechsel sowie auf den Kaliumhaushalt.
Das Hormon ist wesentlicher Faktor folgender Erkrankungen:
- Diabetes mellitus
- Hyperinsulinismus
- Insulinom
- Insulinresistenz
- Metabolisches Syndrom

Das im Blut zirkulierende Insulin entfaltet seine Wirkung durch Bindung an Insulinrezeptoren.
Insulinrezeptor
Letztlich löst die Bindung dieses Hormons an seinen Rezeptor eine Reihe von Kinase-Kaskaden (Kaskade von Phosphorylierungsreaktionen) aus, die durch Signalwege beschrieben werden können.
Diese Signalwege bewirken ein Sinken des Blutglucosespiegels durch
- Förderung der Glucose-Aufnahme (GLUT4-Translokation zur Zelloberfläche)
- Förderung der Glucose-Speicherung (Glykogensynthese) in der Leber und den Muskeln
Dieses Signal wird durch die Aktivierung glucoseverbrauchender Wege unterstützt. Weitere unterstützende Maßnahmen bestehen in der Unterdrückung glucoseliefernder Wege, so zum Beispiel durch Abbau des second messenger cAMP über eine Phosphodiesterase.
Glukoseaufnahme im Muskelgewebe
Das Hormon erhöht in der Muskulatur und im Fettgewebe die Permeabilität (Durchlässigkeit) der Zellmembran für Glucose. Dabei ist zu beachten, dass nicht die Membran selbst permeabler wird, sondern dass vermehrt Carrier-Proteine für Glucose aktiviert werden. Dieses Carrier-Protein ist GLUT4, ein hochaffiner, insulinabhängiger Glucose-Transporter, welcher die Glucose durch erleichterte Diffusion (passiver Transport) in die Zelle transportiert. Folgende physikalische Eigenschaften sind für GLUT4 relevant: sättigbar, nicht aktivierbar oder inaktivierbar, also Regulation nur durch insulinabhängigen Einbau oder Ausbau.
Glukoseaufnahme und Stoffwechsel im Gehirn
Nervenzellen (und Erythrozyten) nehmen Glucose insulinunabhängig auf. Deshalb nehmen die insulinabhängigen Zellen bei einem erhöhten Insulinspiegel mehr Glucose auf, und für die insulinunabhängigen bleibt weniger übrig.
Im Allgemeinen besteht bei Hypoglykämie die Gefahr, dass das auf Glucose angewiesene Nervensystem geschädigt wird. Beachtenswert ist der Effekt, dass Insulin, verabreicht als Nasenspray in direktem Kontakt mit dem ZNS, die Alzheimer-Krankheit positiv beeinflusst.
Auf- und Abbau von Fettgewebe
Das Hormon hemmt die Lipolyse im Fettgewebe und somit den Abbau von Fett. Ein Insulinmangel führt daher zu einer gesteigerten Lipolyse mit Bildung von Ketokörpern und einer daraus resultierenden Ketose.
Förderung des Zellwachstums
Eine weitere zentrale Funktion des Peptidhormons Insulin besteht in der Regulation von Zellwachstum und Proliferation durch die Aktivierung der Transkription von Genen, die für Kontrolle und Ablauf des Zellzyklus von großer Bedeutung sind. Diese Insulinwirkung ist bei Diskursen über Insulinpräparate ein Thema.
Tryptophan-Aufnahme im Gehirn
Höhere Insulinspiegel haben einen leicht steigernden Einfluss auf die Aufnahme von Tryptophan ins Gehirn.
Insulin und die Regelung des Blutzuckerspiegels
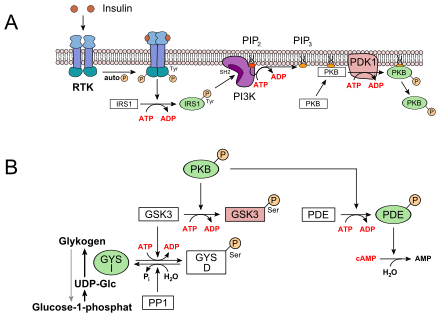
Eine der wichtigsten biologischen Wirkungen des Insulins ist die rasche Beschleunigung der Glucoseaufnahme in Muskel- und Fettzellen und Regulierung der Zwischenspeicherung in der Leber im Rahmen der Regelung des Blutzuckerspiegels:
- In der Leber und der Muskulatur werden die mit der Nahrung aufgenommenen Kohlenhydrate als Glykogen gespeichert. Dies hat ein Absinken der Glucosekonzentration im Blut zur Folge. Die Glucoseaufnahme in die Leberzellen erfolgt insulinunabhängig über GLUT2. Durch Insulin wird eine Rezeptor-Tyrosinkinase (RTK) aktiviert, die eine Signaltransduktion in Gang setzt. Beteiligt sind dabei das Insulinrezeptorsubstrat 1 (IRS1), die Phosphoinositid-3-Kinase (PI3K), der second messenger Phosphatidylinositol-4,5-bisphosphat (PIP2), die Phosphoinositid-abhängige Kinase-1 (PDK1) und schließlich die Proteinkinase B (PKB) (siehe Bild, A). PKB phosphoryliert die Glykogensynthase-Kinase 3, GSK3, die dadurch inaktiviert wird. GSK3 ist eine Kinase, die die Glykogensynthase phosphoryliert und damit inaktiviert (GYS b). GSK3 steht in Konkurrenz zu einer Phosphatase, der Protein-Phosphatase 1 (PP1). Dadurch, dass GSK3 nicht mehr wirken kann, liegt daher immer mehr Glykogensynthase in seiner dephosphorylierten Form vor (GYS a, siehe unteres Bild, B). Außerdem aktiviert die PKB eine Phosphodiesterase, PDE, die cAMP zu AMP hydrolysiert. Infolgedessen erlischt zusätzlich der Signalweg für die Proteinkinase A, die für den Abbau von Glykogen sorgt.
- In der Leber, dem Fettgewebe und der Muskulatur wird unter Insulineinfluss die Triglyceridsynthese stimuliert. Substrate dafür sind neben den Kohlenhydraten mit der Nahrung aufgenommene Lipide.
- In den drei genannten Geweben werden Aminosäuren verstärkt aufgenommen und für die Proteinsynthese verwendet.
Die metabolischen und mitogenen Effekte von Insulin werden über die Bindung an dessen Rezeptor auf der Zelloberfläche der Zielgewebe Leber, Muskel und Fett initiiert.
- Insulin induziert weiterhin die Glykogensynthese und -speicherung in Leber und Muskel, die Triglyceridsynthese in Leber und Fettgewebe sowie die Speicherung von Aminosäuren im Muskel.
- Gleichzeitig hemmt Insulin die hepatische Gluconeogenese und zählt daher insgesamt zu den wichtigsten Regulatoren des Glucosemetabolismus.
Gegenspieler
Fällt der Blutzuckerspiegel im Körper unter einen Wert von 80 mg/dl ab, wird die Insulinproduktion bereits stark reduziert.
Sinkt der Blutzucker weiter ab, treten verschiedene Gegenspieler des Insulins auf:
Die Spiegel dieser gegenregulierenden Hormone steigen bereits deutlich an, wenn der Blutzucker unter 60 mg/dl absinkt.
Beim Typ 1-Diabetes ist oft auch der Gegenregulationmechanismus gestört, was zu zusätzlichen Problemen mit Hypoglykämien führt.
Somatostatin hat einen hemmenden Einfluss auf die Sekretion von Insulin und Glucagon, da er als allgemeiner Hemmer im Körper fungiert.
Vorkommen
Insulinsequenzen von mehr als 100 verschiedenen Spezies sind bekannt. Die Proteinsequenzen der jeweiligen Insuline sind sich ähnlich – sie zeigen Sequenzhomologie – sind aber nicht identisch.
Inwieweit Gene den Glukosestoffwechsel und die damit verbundene Wirkung von Insulin beeinflussen, ist noch nicht zur Gänze geklärt.
Bildung, Speicherung, Freisetzung und Regulierung
Biosynthese
Die Synthese des Hormons erfolgt in den β-Zellen der Langerhansschen Inseln der Bauchspeicheldrüse. Die genetische Information wird von nur einem Genlocus im kurzen Arm des Chromosom 11 codiert. Das Gen besteht aus ungefähr 300 Nukleotiden.
Die mRNA wird zunächst an Ribosomen, die sich auf dem rauen endoplasmatischen Retikulum (ER) befinden, in das Präproinsulin translatiert, das aus 110 Aminosäuren besteht.
Die weitere Prozessierung erfolgt in zwei Schritten, nach der Faltung des Moleküls durch Bildung von Disulfidbrücken entsteht durch Abspaltung von Signalpeptid und C-Peptid das Insulinmolekül.
|
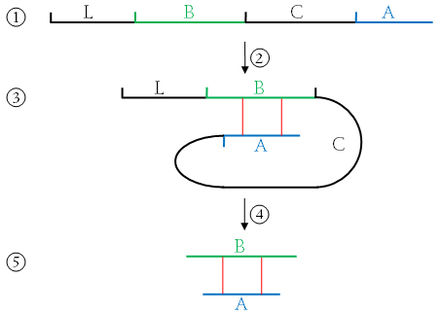
Prozessierung des
Präproinsulin zum Insulin 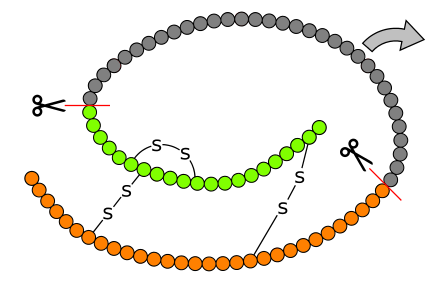
Schematische
Darstellung (Topologiemodell) von Proinsulin (B-Kette orange, A-Kette
grün, C-Peptid grau). Das C-Peptid wird herausgeschnitten. Das
Signalpeptid des Präproinsulins ist nicht
dargestellt. |
Speicherung
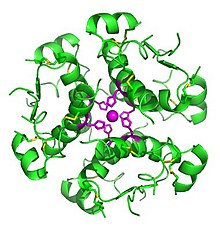
Die Insulinmoleküle werden in den Vesikeln des Golgi-Apparats, der an der Zellmembran der β-Zelle liegt, durch Zink-Ionen zu Hexameren gebunden und so stabilisiert gespeichert (Zink-Insulin-Komplex).
Die hohe Bindungsfreudigkeit von Insulinmolekülen an Zink hat mehrere wichtige Auswirkungen. Insulin ist in der Form von Hexameren und nach dem Zerfall in Dimere noch nicht wirksam, sondern nur als Einzelmolekül. Diese Eigenschaft spielt bei den Insulinpräparaten eine wichtige Rolle. Bei schnellwirkenden Insulinpräparaten ist der zu langsame Zerfall der Molekülverbände unerwünscht und es wird nach Möglichkeiten gesucht den Zerfall zu beschleunigen. Bei langwirkenden Insulinpräparaten wird die Zinkbindung zur Verlängerung der Wirkdauer durch hohe Zinkkonzentrationen gezielt verstärkt. Bei der Entwicklung von oralen Insulinpräparaten wird die Zinkbindung zur Kopplung von Insulin an Transportmoleküle genutzt.
Ausschüttung

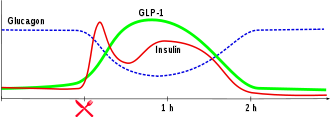
Die Ausschüttung von Insulin ins Blut erfolgt durch Exozytose.
Die Insulinausschüttung erfolgt oszillierend. Alle drei bis sechs Minuten wird Insulin in die Blutbahn abgegeben. Nach der Nahrungsaufnahme ist bei Stoffwechselgesunden ein biphasischer Verlauf der Insulinsekretion feststellbar: Der erste „Insulinpeak“ hat seine Spitze nach drei bis fünf Minuten und dauert zehn Minuten. Danach schließt sich eine zweite Phase an, die so lange anhält, wie eine Hyperglykämie besteht. Die erste Phase besteht aus den gespeicherten Insulinmolekülen, die zweite Phase vor allem aus neu gebildetem Insulin.
Das C-Peptid wird erst bei Anstieg des Blutzuckerspiegels aus dem Proinsulin durch Peptidasen herausgeschnitten und gemeinsam mit dem aktiven Insulin und Zink ausgeschüttet. Durch den Nachweis von C-Peptid im Serum kann die endogene Insulinproduktion gemessen werden. So kann bei Diabetikern eine Aussage darüber getroffen werden, wie viel Insulin noch vom Körper selbst hergestellt wird, da das synthetische Produkt keine C-Sequenz enthält.
Regulierung
Die Hauptaufgabe von Insulin besteht darin, im Zusammenwirken mit Glukagon den Blutglukosespiegel ständig zwischen bestimmten Werten zu halten. Gerät der Blutzuckerspiegel unter einen gewissen Wert droht das hypoglykämische Koma und der Tod, gerät er darüber, drohen Schäden an Gefäßen, Nierenkörperchen und anderen Geweben. Zusammen mit Leptin scheint Insulin aber auch maßgeblichen Anteil an der Energiehomöostase zu haben. Dabei wirkt aber nicht nur Leptin auf Insulin ein, sondern das Insulin wirkt selbst ebenfalls auf das Leptin ein, genau auf die Aufnahme von Leptin im Gehirn.
Aufgrund der lebenswichtigen Funktion muss die biochemische Implementierung des Regelkreises robust sein. Da Zellen nicht „denken“ können, kann es nur so sein, dass Einzelzellen wie ein Zustandsautomat funktionieren bzw. dass wenige Zellen dermaßen zusammenwirken, dass ein Reiz sinnvoll verrechnet wird.
Der Hauptreiz zur Ausschüttung von Insulin aus der β-Zelle ist der Blutzuckerspiegel (ab 5 mmol Glucose/l Blut), und dieser wird direkt von der β-Zelle „gemessen“. Biochemisch umgesetzt wird das mit Hilfe von ATP-gesteuerten Kaliumkanälen, etc., siehe Glukosegesteuerter Freisetzungsmechanismus.
Die Hormone Gastrin, Sekretin, GIP und GLP-1 modifizieren den grundlegenden Glukose-Insulin-Regelkreis indem sie auf die β-Zelle wirken.
GIP wird von K-Zellen, welche sich in der Schleimhaut des Duodenums befinden und die Glukose im Chymus „messen“, ins Blut sezerniert und erhöht die Insulinausschüttung der β-Zellen. Das GIP wirkt also auf die β-Zellen zu einem Zeitpunkt ein, wo die Glukose aus der Nahrung noch nicht ins Blut gelangt ist.
GLP-1 wird von L-Zellen, welche sich in der Schleimhaut des Ileum und des Caecum befinden und die Glukose im Chymus „messen“, ins Blut sezerniert und erhöht ebenfalls die Insulinausschüttung der β-Zellen. Der Großteil der Nährstoffe wird bereits im Jejunum extrahiert und ins Blut bzw. in die Lymphe abgegeben. Im Caecum findet keinerlei Absorption von Nährstoffen, sondern hauptsächlich eine Fermentation statt.
Zusätzlich sind β-Zellen sowohl vom parasympathischen als auch sympathische Nervensystem innerviert:
Das parasympathische NS wird bei Leptin-Insuffizienz (oder auch bei Leptin-Resistenz) aktiviert und wirkt allgemein trophotropisch, d.h., es bewirkt die Speicherung von Energie. Es erhöht also die Insulinabgabe. Zumindest bei Mäusen wirkt es außerdem gleichzeitig auf Adipozyten ein und erhöht ihre Empfindlichkeit für Insulin, während die Empfindlichkeit der Leber- und Muskelzellen nicht beeinflusst wird. Glukose wird in Leber- und Muskelzellen zu Glykogen verbaut und derart gespeichert, in den Adipozyten erfolgt De-novo-Lipogenese und Speicherung von TGs.
Das sympathische NS wird bei Leptin-Suffizienz aktiviert und wirkt allgemein ergotropisch, d.h., es bewirkt die Erhöhung des Energieverbrauchs. Es senkt die Insulinabgabe. Die Senkung der Insulinabgabe scheint einem erhöhten Energieverbrauch entgegenzuwirken, denn die Muskelzellen können sehr viel mehr Glukose aus dem Blut aufnehmen, wenn sie möglichst viele GLUT-4 Transporter auf der Oberfläche aufweisen. Sie müssen also auf die bereits gespeicherte Energie in Form von Glykogen und Fettsäuren zurückgreifen.
Glukosegesteuerter Freisetzungsmechanismus
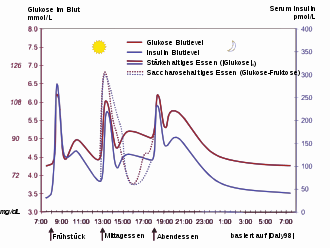
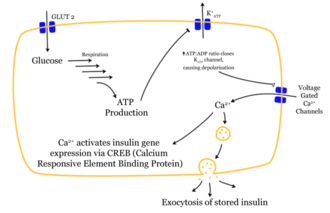
Das Eindringen eines Glukose-Moleküls in die β-Zelle setzt eine Wirkungskette in Gang. Nachdem die Glukose durch den GLUT1-Transporter in die Zelle gelangt ist, wird sie durch Glykolyse verstoffwechselt. Das dabei entstehende ATP hemmt den Ausstrom von Kalium-Ionen (ATP-sensitive Kaliumkanäle). So kommt es durch den stark verminderten Kaliumausstrom zur Depolarisation, weil die Stabilität des Membranpotentials nicht weiter durch Kaliumausstrom erhalten wird. Das depolarisierte Membranpotential bewirkt eine Öffnung spannungsabhängiger Kalzium-Kanäle. Der Einstrom der Kalziumionen ist der entscheidende Reiz für die Verschmelzung der insulinhaltigen Vesikel mit der Zellmembran.
Die gespeicherten Insulinmoleküle werden durch Verschmelzen der Membranen (Exozytose) aus den β-Zellen in den Extrazellularraum und weiter in den Blutkreislauf freigesetzt. Dabei werden die Speicher-Hexamere aufgetrennt. Der Insulinspiegel im Blut steigt an.
Halbwertszeit und Abbau
Die biologische Halbwertszeit einzelner Insulinmoleküle im Blutkreislauf liegt bei circa fünf Minuten.
Das Insulin wird über manche Insulinrezeptoren in die Zellen aufgenommen, dort abgebaut und somit verbraucht. In der Leber und in der Niere wird Insulin durch Insulinase inaktiviert, die Insulinase oder genauer Glutathion-Insulin-Transhydrogenase spaltet die Disulfidbrücken zwischen der A- und der B-Kette auf, wodurch das Insulin in zwei Teile zerfällt und wirkungslos wird. Die Abbauprodukte werden durch die Niere ausgeschieden, ebenso 1,5 % des noch intakten Insulins. Die kurze Zeitspanne der Aktivität des Insulins zeigt, dass die physiologische Steuerung des Zuckerstoffwechsels im gesunden Körper sehr schnell funktioniert; diese Geschwindigkeit kann bei der Behandlung des Diabetes mellitus praktisch nicht erreicht werden.
Insulin als Arzneistoff
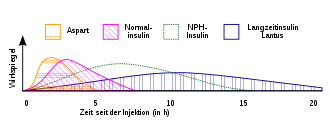
In der Insulintherapie werden verschiedene Insulinpräparate verwendet. Die häufigste und älteste Verabreichungsart ist die subkutane Injektion. Für diesen Zweck steht eine Reihe von kurz-, mittel- und langwirkenden Humaninsulinen und Insulinanaloga zur Verfügung. Werden diese zur Therapie kombiniert, so ist auf die unterschiedliche Halbwertzeit besonders zu achten.
- sehr schnell und kurz wirkend: Insulin glulisin, Insulin lispro, Insulin aspart
- kurz wirkend: Normalinsulin (= gelöstes Humaninsulin)
- intermediär wirkend: NPH-Insulin, biphasisches Insulin lispro, biphasisches Insulin aspart
- lang wirkend: Insulin detemir, Insulin glargin, Insulin degludec
Peroral ist Insulin unwirksam, da die Eiweißketten im Magen-Darm-Trakt von körpereigenen Enzymen abgebaut werden, bevor sie ihre Wirkung entfalten können. Untersucht wird, inwieweit sich Insuline in Nanopartikel verkapseln lassen, um auf diesem Wege „unverdaut“ in den Blutkreislauf eingeschleust werden zu können. Jüngere Entwicklungen wie Präparate zur Inhalation, die das Insulin über die Atemwege zuführen, haben sich auf dem Markt bisher nicht behaupten können.
In der Vergangenheit wurde Insulin im Rahmen der Insulinschocktherapie zur Behandlung von Menschen mit psychischen Krankheiten eingesetzt. Diese Methode der Behandlung wurde zum Beispiel im biografischen Film A Beautiful Mind an John Nash praktiziert. Dieses Verfahren wird nicht mehr praktiziert.
Missbrauch von Insulin
Insulin steht auf der Liste der verbotenen Doping-Substanzen, da es zu mehreren Zwecken missbraucht werden kann. Da Insulin der durch Somatropin verringerten Glukoseaufnahme in die Muskelzellen entgegenwirkt, wird es oft zur Kompensation dessen unerwünschter Nebenwirkung verwendet. Andere Anwendungen sind die Förderung der Füllung der Glykogenspeicher bei Ausdauersportlern und die Unterstützung des Aufbaus von Muskelmasse.
Die missbräuchliche Selbstverabreichung von Insulin, um den Blutzuckerspiegel übermäßig abzusenken, führt zum Krankheitsbild der Hypoglycaemia factitia.
Im März 2008 wurde der Krankenpfleger Colin Norris in Newcastle zu einer Freiheitsstrafe von 30 Jahren verurteilt, weil er vier seiner Patientinnen durch Injektion zu hoher Insulindosen ermordet hatte.Colin Norris, 'Angel of Death' nurse, convicted. In: The Telegraph. 2. März 2008


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 26.04. 2026