Taxon
Taxon ( das, Pl.: Taxa; von altgriechisch τάξις táxis, (An-)Ordnung, Rang) bezeichnet in der Systematik der Biologie eine Einheit, der entsprechend bestimmter Kriterien eine Gruppe von Lebewesen zugeordnet wird. Meist drückt sich diese Systematik durch einen eigenen Namen für diese Gruppe aus.
Grundsätzlich lassen sich Formtaxa und echte Taxa unterscheiden.
Formtaxa weisen ähnliche Merkmale auf, müssen jedoch stammesgeschichtlich nicht direkt miteinander verwandt sein (etwa Wirbellose, Laubbäume oder Einzeller). Die entsprechenden Merkmale sind an verschiedenen Orten unabhängig voneinander entstanden (analoge Entwicklung).
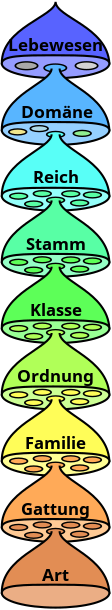
Echte Taxa hingegen bilden idealerweise geschlossene Abstammungsgemeinschaften („natürliche Gruppen“), sodass die entsprechende Systematik ein Abbild der evolutionär entstandenen Verwandtschaftsbeziehungen liefert. Die Verwandtschaftsbeziehungen weisen eine hierarchische, mehrfach-verzweigte Baumstruktur („Stammbaum“) auf. Entsprechend hierarchisch ist auch die biologische Systematik aufgebaut: Die meisten Taxa sind Subtaxa eines übergeordneten Taxons und enthalten selbst Subtaxa. In der traditionellen Systematik sind die Hierarchieebenen, die auch als Rangstufen oder Ränge bezeichnet werden, benannt. Die höchste Rangstufe ist die Domäne (enthält die Taxa Bakterien, Archaeen und Eukaryoten). Die nächstniedrigere Rangstufe ist das Phylum (innerhalb der Bakterien und Archaeen) oder das Reich (innerhalb der Eukaryoten), die wiederum jeweils in niedrigere Rangstufen gegliedert werden. Bei den Eukaryoten werden diese weiteren Rangstufen in absteigender Reihenfolge traditionell als Stämme, Klassen, Ordnungen, Familien, Gattungen und Arten bezeichnet. Diese Rangstufen werden zudem oft in Über- und/oder Unterränge gegliedert (z.B. Überordnungen oder Untergattungen). Die eigentlichen Taxa sind die individuell benannten Einheiten innerhalb einer Rangstufe. Mit wachsender Bedeutung der Kladistik in der Biologie, aus deren Methoden zur Ermittlung der Verwandtschaftsbeziehungen sich, im Vergleich zur traditionellen Systematik, ein Vielfaches an potenziellen Hierarchieebenen ergibt, wird auf die Benutzung der traditionellen Rangstufen zunehmend verzichtet.
Da Viren nicht als Lebewesen gelten, steht die Virus-Taxonomie außerhalb der hier behandelten Hierarchie.
Die Verwandtschaft der jeweils zu einem übergeordneten Taxon zusammengefassten Taxa kommt durch mindestens ein gemeinsames Merkmal zum Ausdruck, durch das sich diese Taxa von anderen gleichrangigen Taxa unterscheiden. In der kladistisch geprägten Systematik haben diese Merkmale per Definition jeweils einen gemeinsamen evolutionären Ursprung, das heißt, sie sind einander homolog und wurden von einer Stammart ererbt. Während in der Kladistik das Taxon, das durch eine bestimmte Stammart definiert wird, immer auch alle deren Nachfahren (also sämtliche Subtaxa aller Rangstufen) einschließt, gibt es in der traditionellen Systematik auch Taxa, die nicht alle Nachfahren der Stammart enthalten. So enthält beispielsweise die Klasse der Reptilien nicht die Vögel (sondern diese bilden eine eigene Klasse der Landwirbeltiere), obwohl die Vögel nach weit verbreiteter Auffassung die engsten lebenden Verwandten der Krokodile (einer traditionellen Ordnung der Reptilien) sind.
Die Regeln für die Vergabe wissenschaftlicher Namen für Taxa sind Inhalt des jeweils zuständigen nomenklatorischen Regelwerkes (ICZN, ICBN, ICNB). Die Einordnung nach dem Grad der Verwandtschaft in das entsprechende hierarchische System wird von der akademischen Disziplin der sich auf die biologische Systematik stützenden Taxonomie vorgenommen. Unterschiedliche methodische Ansätze und/oder Merkmalsinterpretationen können zu alternativen Ergebnissen bei der Ermittlung der Verwandtschaftsverhältnisse führen und mithin zu voneinander abweichenden Taxon-Konzepten und oft auch zu unterschiedlich benannten Taxa. Traditionell werden häufig aus dem Lateinischen oder Griechischen abgeleitete Namen vergeben. Ein solchermaßen mit Namen versehenes Taxon wird als nominelles oder formelles Taxon bezeichnet.
Formtaxa
Formtaxa sind Gruppen von Lebewesen, die von der Systematik zwar als nicht stammesgeschichtlich zusammengehörig erkannt worden sind, aber aus praktischen Gründen dennoch mit eigenem Namen beibehalten werden. Beispiele für solche Formtaxa sind:
- die Wirbellosen (Invertebrata), die sich dadurch auszeichnen, dass sie keine Knochen besitzen. Zu dieser von Jean-Baptiste de Lamarck benannten Gruppe gehört der weitaus größte Teil aller Tierarten. Wirbellose Tiere wie Schwämme (Porifera), Insekten (Insecta) oder Seesterne (Asteroidea) sind jedoch nur sehr entfernt miteinander verwandt und bilden keine natürliche Verwandtschaftsgruppe.
- die Flechten (Mycophycophyta), die als symbiotische Lebensgemeinschaften von Pilzen und Algen keine selbständigen Lebewesen sind, sondern aus zwei fundamental verschiedenen Partnern bestehen. Dennoch werden sie teils bis heute als eigenes Taxon mit dazugehörigen Untergruppierungen wie Gattungen und Arten geführt.
- die Fungi imperfecti (Deuteromycota), die eine Sammelgruppe für all diejenigen Pilze darstellen, die entweder über keine sexuellen Fortpflanzungsstrukturen verfügen oder deren Fortpflanzungswege noch unverstanden sind. Teilweise werden sogar ungeschlechtliche Stadien von Pilzen aufgenommen, die ansonsten durchaus zur sexuellen Vermehrung im Stande sind. Stammesgeschichtlich gehören viele Organismen dieser Gruppe zu anderen Gruppen der Pilze wie den Schlauch- (Ascomycota) oder Ständerpilzen (Basidiomycota). Hierbei werden Anamorphe und Teleomorphe nicht selten unter verschiedenen Taxa geführt.
- die Protisten (Protista), die als Gesamtheit aller Organismen mit echtem Zellkern (Eukaryota) definiert werden, ohne Tiere (Animalia), Pilze (Fungi) oder Pflanzen (Plantae) zu sein. Oft werden sie Einzeller genannt, obwohl sich in dieser Gruppe zahlreiche mehrzellige Organismen finden, die, wie beispielsweise die Braunalgen (Phaeophyta), zu den größten Lebewesen unseres Planeten zählen.
Während heute weitgehend unumstritten ist, dass es sich bei vorstehenden Beispielen um Formtaxa handelt, ist die Frage, was als echtes Taxon anzusehen ist, problematisch und hängt von grundsätzlichen Überlegungen hinsichtlich der Systematik ab: Was aus Sicht der einen ein gültiges Taxon ist, kann aus der Perspektive einer anderen lediglich eine formelle Gruppe sein.
Synonyme
Am einfachsten aufzulösen sind Unstimmigkeiten, die lediglich aufgrund synonymer Benennungen entstehen. So können zum Beispiel zwei Biologen, die in unterschiedlichen Regionen derselben Art begegnen, dieser zunächst unterschiedliche Namen zuordnen oder es können zwei ursprünglich als getrennt angesehene Arten als Teilpopulationen einer einzigen erkannt werden. Auf diese Weise hat zum Beispiel der Feldhase (Lepus europaeus) im Laufe der Zeit dreiundvierzig verschiedene wissenschaftliche Bezeichnungen (Synonyme) erhalten. Eine andere Möglichkeit ist, dass aufgrund neuer wissenschaftlicher Erkenntnisse eine veraltet geglaubte Gruppierung doch wieder als echtes Taxon angesehen wird, aber in Unkenntnis einer vielleicht Jahrhunderte zurückliegenden Benennung zunächst einen neuen Namen erhält. In beiden Fällen wird der Konflikt dadurch gelöst, dass der älteste verfügbare Name als „gültiger“ Name nach der Prioritätsregel Vorrang über die „jüngeren Synonyme“ erhält.
Nur in seltenen Fällen gibt es Ausnahmen, z.B. geht Equus ferus Boddaert, 1785 vor Equus caballus Linnaeus 1758, da nach einer Entscheidung der ICZN-Kommission von 2003 dem jüngeren Namen Vorrang vor dem älteren gegeben wurde. Eine andere Ausnahme ist Tyrannosaurus rex Osborn, 1905, ausnahmsweise mit Vorrang vor Manospondylus gigas Cope, 1892.
Der umgekehrte Fall, dass zwei verschiedenen Taxa derselbe Name zugeordnet wird, kann ebenfalls auftreten. Auch hier ist dann der Name nur für das zuerst benannte Taxon verwendbar (wenn überhaupt), für das andere Taxon kann dieser Name nicht verwendet werden.
Von größerer theoretischer Bedeutung für die Gültigkeit eines Taxons sind aber tiefergehende systematische Überlegungen zu der Frage, nach welchen Kriterien bestimmt wird, welche Gruppe von Lebewesen ein echtes Taxon bildet und welche nicht. Als bedeutendste Systeme gelten zu Beginn des 21. Jahrhunderts die klassische und die kladistische Taxonomie. Ihre folgende Gegenüberstellung ist zur Herausarbeitung der Unterschiede idealisiert; in der biologischen Praxis werden oft beide Systeme nebeneinander oder in Kombination verwendet.
Taxa in der klassischen Taxonomie
Die klassische Taxonomie geht auf ein durch den schwedischen Naturforscher Carl von Linné (dessen Name in dieser Zeit Carl Linnæus lautete) eingeführtes Klassifikationssystem zurück. Danach werden nicht nur alle Lebewesen in eine Hierarchie ineinander verschachtelter Taxa gruppiert, sondern diesen Gruppen wird auch je eine Kategoriestufe zugeordnet. Die basale Kategorie, die Grundlage für die gesamte Klassifikation, bildet die Art, die nach den unterschiedlichsten Gesichtspunkten definiert sein kann, bei sich geschlechtlich fortpflanzenden Organismen aber oft durch das Kriterium der Interfertilität festgelegt ist, was bedeutet, dass Mitglieder einer Art unter natürlichen Bedingungen gemeinsame fruchtbare Nachkommen zeugen können. Diese Definition des Artbegriffs ist wie auch ihre Alternativen nicht unproblematisch, eine weitergehende Erläuterung findet sich im Artikel zur Art.
Namensregeln für klassische Taxa
Jede Art (Spezies) erhält nach einer auf Linné zurückgehenden Konvention einen zweiteiligen (binären) Namen, der aus dem Namen der nächsthöheren Kategorie, der Gattung, und dem so genannten Artepithet oder Artepitheton (in der Zoologie: dem Artnamen) zusammengesetzt ist und deshalb auch als Binomen bezeichnet wird. Diese beiden Teile des Art-Binomens werden nach den Konventionen heute meist kursiv geschrieben: Gattungsname groß und Artepithet/-name klein. Alle Taxa höheren Ranges haben nach diesen Regeln einen aus einem einzelnen Wort bestehenden Namen, der mit einem Großbuchstaben anfängt und nicht kursiv geschrieben wird. Jeder Gattungsname kann innerhalb des Organismenreiches nur einmal vergeben werden (derselbe Gattungsname kann aber in der Zoologie und in der Botanik parallel vergeben werden). Der zweite Teil des Art-Binomens kann in verschiedenen Gattungen mehrmals vergeben und verwendet werden.
Linné führte neben der Gattung auch die Ränge Ordnung, Klasse und Reich ein. Später wurde dieses System um die zwischen Gattung und Ordnung gelegene Familie und den zwischen Klasse und Reich gelegenen Stamm (in der Zoologie) beziehungsweise die Abteilung (in der Botanik) erweitert. In neuester Zeit wurden noch weitere Ränge wie Domäne, Reihe, Kohorte, Legion oder Tribus eingeführt. Alle Kategorien können noch durch die Vorsätze „Über-“ beziehungsweise „Unter-“ feiner untergliedert werden. Bei manchen Taxa lässt sich schon durch den Namen feststellen, welcher Kategorie der klassischen Rangsysteme sie angehören: So bezeichnet zum Beispiel die Endung „-aceae“ eine Familie von Pflanzen, während eine Familie von Tieren die Endung „-idae“ trägt. Die genauen Benennungsregeln sind in den jeweiligen und voneinander unabhängigen Regelwerken der Biologischen Nomenklatur festgeschrieben.
Für die Benennung von Unterarten werden dreiteilige (trinominale) Namen verwendet. Sie bezeichnen Taxa innerhalb einer Art, die zwar gut charakterisiert und voneinander unterschieden werden können, deren Individuen aber mit denen anderer, ebenfalls gut abgrenzbarer Taxa innerhalb der gleichen Art noch fruchtbar kreuzungsfähig sind. Beim Trinomen von Unterarten wird dem Art-Binomen der Unterartname, stets kleingeschrieben, nachgestellt, und genau wie das Art-Binomen wird das Unterart-Trinomen vollständig kursiv geschrieben.
Abgrenzung klassischer Taxa
Die Abgrenzung der Taxa erfolgte bei Linné noch nach rein formellen Kriterien, den so genannten essentiellen Merkmalen. So unterteilte er zum Beispiel die Blütenpflanzen nach der Zahl und Anordnung der Staub- und Fruchtblätter der Blüte, also nach ihren Fortpflanzungsstrukturen. Schon sein Zeitgenosse, der französische Naturforscher Georges Louis Leclerc Graf von Buffon, wies jedoch auf die Willkürlichkeit dieses Kriteriums hin.
Seit den bahnbrechenden Arbeiten des britischen Geologen und Naturforschers Charles Darwin hat sich stattdessen die Vorstellung durchgesetzt, dass die Einteilung der Lebewesen in Taxa ihre natürlichen Verwandtschaftsverhältnisse berücksichtigen sollte. Aber auch diese Sicht- und Vorgehensweise war nicht neu. Viel frühere Forscher einschließlich Linné hatten immer wieder, sobald neue Erkenntnisse über die Verwandtschaften bekannt geworden waren, die Taxonomie angeglichen und dementsprechend die Fledermäuse nicht mehr den Vögeln und die Wale nicht mehr den Fischen zugeordnet. Ausgesprochen willkürliche Taxa, die auf rein formellen Kriterien basierten, wie beispielsweise Flugtiere, wurden spätestens seit 1700 nicht mehr verwendet und flossen in Linnés moderne wissenschaftliche Namensgebung nicht mehr ein.
Allerdings versuchen klassische Systematiken, wie die von dem deutsch-amerikanischen Evolutionsbiologen Ernst Mayr und dem amerikanischen Paläontologen George Gaylord Simpson formulierte so genannte evolutionäre Systematik nicht nur die stammesgeschichtlichen Verzweigungen in die Definition von Taxa einfließen zu lassen, sondern auch weitere Kriterien zur Klassifikation heranzuziehen, um wesentliche evolutionsgeschichtliche Neuerungen abzubilden.
- So stellt Ernst Mayr zum Beispiel in seiner 1990 vorgestellten Klassifikation die einzelligen Protisten (Protista) in einer eigenen Unterdomäne den Mehrzellern (Metabionta) gegenüber, zu denen er Tiere (Animalia), Pflanzen (Plantae) und Pilze (Fungi) zählt, obwohl das Kriterium der Mehrzelligkeit im Laufe der Evolutionsgeschichte sehr wahrscheinlich mehrfach unabhängig voneinander entstanden ist. Entscheidend ist nach Mayr hier aber nicht der nur molekulargenetisch feststellbare Verwandtschaftsgrad, sondern eben auch die äußere Erscheinungsform (Morphologie).
- Das System der bedeutenden Evolutionsbiologin Lynn Margulis (1988, 1996) fasst bewusst entgegen den (wahrscheinlichen) Verwandtschaftsverhältnissen die beiden Gruppen der Archaebakterien (dann Archaebacteria) und echten Bakterien (dann Eubacteria) zu dem Taxon der Prokaryoten (Procaryota) zusammen, weil sie als Organismen, deren Zellen keinen Zellkern besitzen, eine andere Organisationsform haben als die Eukaryoten (Eukaryota), in deren Zellen sich ein solcher befindet. Der Strukturunterschied, der mit der „Erfindung“ des Zellkerns einherging, wird damit als bedeutender eingeschätzt als eine möglichst genaue Abbildung des evolutionären Stammbaums.
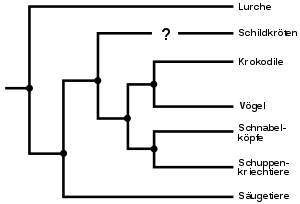
- Ein weiteres Beispiel lässt sich an den beiden Gruppen der Vögel (Aves) und Reptilien (Reptilia) erkennen. Sie werden in der klassischen Taxonomie als gleichrangige Klassen geführt, obwohl heute unumstritten ist, dass erstere entwicklungsgeschichtlich aus letzteren hervorgegangen sind, was sich auch darin äußert, dass eine Kriechtiergruppe, die Krokodile (Crocodylia), stammesgeschichtlich enger mit den Vögeln verwandt ist als mit anderen Kriechtieren wie zum Beispiel den Schlangen (Serpentes). Die Abtrennung der Vögel als separate Klasse wird jedoch von traditionellen Taxonomen wie Mayr oder Simpson nicht nur mit Verweis auf die bedeutenden Unterschiede in Anatomie und Physiologie, die Vögel heute von den Reptilien trennen, sondern auch in Hinsicht auf den bedeutenden ökologischen Rollenwechsel, der mit der Eroberung des Luftraums verbunden war, als gerechtfertigt angesehen.
- Die Robben (Pinnipedia), die stammesgeschichtlich von landlebenden Raubtieren (Carnivora) abstammen, aber wegen des extremen Wandels ihres Lebensraumes oft als selbständige Ordnung im gleichen Rang wie die Raubtiere geführt werden, sind ein vergleichbarer Fall.

Charakteristisch ist in jedem Fall, dass nicht nur das Kriterium der stammesgeschichtlichen Verwandtheit, sondern darüber hinaus auch deutliche morphologische Unterschiede (Diskontinuitäten), ökologische Nischenwechsel, die Komplexität des anatomischen Bauplans oder die Artenvielfalt einer Gruppe zur Abgrenzung eines Taxons herangezogen werden. Daraus folgt, dass Mitglieder eines Taxons durch ihre weitere evolutionäre Entwicklung in ein anderes, gleichrangiges Taxon übertreten können, wie dies zum Beispiel bei den Vögeln geschehen ist, die wie bereits erwähnt nach klassischer Sicht eine gleichrangige Gruppe neben den Kriechtieren bilden. Umgekehrt werden nach rein stammesgeschichtlichen Kriterien gebildete Gruppen, die morphologisch stark voneinander abweichende Untertaxa enthalten, aus klassischer Sicht oft abgelehnt; so wird zum Beispiel die Zusammenfassung der Schwestergruppen der Vögel und Krokodile zu den Archosauriern (Archosauria) als absurd verworfen.
Während heutige Anhänger der klassischen Systematik polyphyletische Taxa, also Gruppen, die nicht einmal ihren letzten gemeinsamen Vorfahren einschließen, dennoch meist ablehnen, besteht weitgehende Einigkeit, dass paraphyletische Taxa, Gruppen, die nicht alle Nachkommen ihres letzten gemeinsamen Vorfahren enthalten, nicht nur erlaubt, sondern wegen der – trotz neu eingeführter Stufen wie Kohorte oder Legion – begrenzten Zahl an Kategorien nahezu unumgänglich sind.
Als Vorteil der klassischen Taxon-Definition gilt ihre vergleichsweise Stabilität: Da sich wissenschaftliche Ansichten über die genauen Verwandtschaftsverhältnisse von Lebewesen weitaus schneller wandeln können als Ansichten über die äußere Erscheinungsform, ist eine solche Klassifikation insbesondere für nicht-biologische Anwendungsgebiete wie zum Beispiel in der Land- und Forstwirtschaft von größerer Praxisrelevanz. Als Nachteil gilt jedoch heute die relative Willkürlichkeit bei der Abgrenzung der Taxa: So ist in einem gegebenen Fall ohne genaue Angaben des Taxonomen für Außenstehende oft nicht zu erkennen, welches Kriterium – Diversität, ökologischer Nischenwechsel oder stammesgeschichtliche Verwandtschaft – gerade zur Abgrenzung des Taxons herangezogen wurde. Kritiker der klassischen Taxonomie haben daher in Anlehnung an den englischen Biologen Thomas Henry Huxley ironisch die Einführung eines eigenen Reiches „Psychozoa“ für den Menschen gefordert, das gleichrangig neben den Reichen der Tiere, Pflanzen und Pilze stehen soll, um den bedeutenden ökologischen Nischenwechsel des Menschen beim Verlassen der afrikanischen Savanne auch taxonomisch zur Geltung zu bringen.
Taxa in der kladistischen Taxonomie
Einen gänzlich anderen Weg bei der Abgrenzung von Taxa geht die auf den deutschen Systematiker und Insektenforscher Willi Hennig zurückgehende so genannte kladistische Taxonomie, die sich auf die Ergebnisse der kladistischen Systematik stützt. Ihr Grundprinzip ist, dass nur natürliche Gruppen von Lebewesen benannt werden sollten. Als solche werden nur Fortpflanzungs- und Abstammungsgemeinschaften akzeptiert, also Gruppen von Lebewesen, die einen gemeinsamen Genpool bilden (Arten) oder alle Nachkommen eines gemeinsamen Vorfahren enthalten. Diese monophyletische Taxa werden auch Kladen (Plural, von Singular die Klade) oder Monophyla genannt. Kriterien wie morphologische Diskontinuitäten, Artenvielfalt etc. werden bewusst nicht zur Abgrenzung eines Taxons herangezogen, die Klassifikation soll damit ausschließlich die natürlichen Verwandtschaftsverhältnisse der Lebewesen getreu abbilden. Unter kladistisch orientierten Taxonomen wird mitunter der Begriff Taxon dann als Synonym für Klade gebraucht, da andere Taxa nicht als gültig akzeptiert werden.
Mit der Zurückweisung paraphyletischer Gruppen einher geht die Ablehnung der als Überbleibsel der typologisch orientierten Klassifikation des 18. und 19. Jahrhunderts angesehenen Kategorien. Sie gelten der kladistischen Taxonomie mit Ausnahme der allerdings etwas anders definierten Art als biologisch bedeutungslos, weil in der Natur zwar mitunter eine vertikale, aber keine horizontale Ordnung der Taxa existiert: So ergibt es zwar Sinn, etwa davon zu sprechen, dass die Raubtiere (Carnivora) eine Untergruppe der Säugetiere (Mammalia) sind, die wiederum eine Untergruppe der Wirbeltiere (Vertebrata) bilden. Dagegen wird die Aussage, dass den Entenvögeln (Anatidae) dieselbe Rangstufe zukommt wie den Korbblütlern (Asteraceae), als wissenschaftlich leer betrachtet, die Vergleichbarkeit aus klassischer Sicht gleichrangiger Taxa also bestritten. Um die Tradierung biologischer Informationen nicht zu gefährden, werden die alten Taxonnamen allerdings fast immer beibehalten – ohne die teilweise in diesen Namen enthaltenen Rangstufen zu berücksichtigen.
Die Bedeutung der traditionellen Ränge geht in einem kladistischen System auf die so genannten Schwestergruppen (Adelphotaxa) über, darunter versteht man zwei Taxa, die einst durch den Prozess der Artbildung (Speziation) aus einer Stammart hervorgegangen sind, welche nach der Aufspaltung dann gemäß strenger kladistischer Anschauung als erloschen, also nicht mehr existent gilt. Durch weitere Aufspaltungsprozesse (Kladogenese) können sich aus den beiden ursprünglichen Schwesterarten mit der Zeit zwei nun zahlreiche Arten umfassende Gruppen von Lebewesen entwickeln, die aber zu jedem Zeitpunkt im selben Verwandtschaftsverhältnis – eben als Adelphotaxa – zueinander stehen. Phänomene wie horizontale Genübertragung oder insbesondere bei Pflanzen Hybridbildung können dieses Bild komplizieren, von grundsätzlicherer Bedeutung sind diese Prozesse auf lange Sicht zumindest für Organismen mit echtem Zellkern (Eukaryota) jedoch nicht.
Beispiele für Schwestertaxa sind:
- die Krokodile (Crocodylia) und Vögel (Aves), die als Archosaurier (Archosauria) zusammengefasst werden. Nur moderne Lebewesen sind hierbei berücksichtigt, der Begriff Schwestertaxon ändert sich also, je nachdem, ob auch fossile Arten hinzugezogen werden oder nicht.
- die Strahlenflosser (Actinopterygii) und Muskelflosser (Sarcopterygii), allerdings nur, wenn letztere Gruppe auch die Landwirbeltiere (Tetrapoda) umfasst.
Nicht verwechselt werden darf der Begriff der Schwestertaxa mit den ursprünglichen Unterarten, aus denen diese hervorgegangen sind. Während sich zwei Schwestertaxa im idealen Fall allein aus der Merkmalsanalyse heute existierender Lebewesen ableiten lassen, ist ihre Stammart beziehungsweise die beiden Schwesterarten, in die diese sich aufgeteilt hat, heute auch bei bester fossiler Überlieferung in nahezu keinem Fall mehr rekonstruierbar. Ausnahmen bestehen nach strenger Ansicht nur bei einer wissenschaftlich beobachteten Artbildung – ein äußerst seltenes Phänomen. Aus diesem Grunde werden in kladistischen Taxonomien auch niemals Vorfahren beziehungsweise Nachkommen eines Taxons identifiziert: Alle existierenden (monophyletischen) Taxa einer gegebenen Gruppe, die auch fossile Gruppen umschließen kann, werden in ein System aus Schwestertaxa eingeordnet, von denen jedes zwar als logische Zusammenfassung oder Teilmenge, niemals aber als historischer Vorgänger oder Nachfolger anderer Taxa gelten kann. Dargestellt wird dieses System in diagrammatischer Weise in einem so genannten Kladogramm, einem Baumdiagramm, in dem sich jeder Ast idealerweise jeweils in zwei Unteräste aufspaltet. Am Ende der feinsten Verästelung sitzen die (fossilen oder modernen) Arten; die Knotenpunkte werden dagegen nicht benannt, sie stehen für die virtuellen, das heißt nie festgelegten Stammarten.
Bei der Benennung von Taxa nach kladistischer Systematik ergeben sich gewisse Schwierigkeiten: Wie bereits erwähnt, führen klassische Taxa oft ihre Rangstufe im Namen mit, daneben ist durch die binäre Nomenklatur für Arten festgeschrieben, dass ein Artname Informationen über seine Gattung enthält. Damit ist es erstens problematisch, zwischen Gattung und Art gelegene Taxa zu benennen, zweitens zieht ein Wechsel in der systematischen Stellung einer Art zugleich auch immer eine Umbenennung nach sich. Deshalb gibt es heute Bestrebungen, die bisherige Nomenklatur zugunsten von neuen auf die kladistische Taxonomie zugeschnittenen Namensregeln zu überwinden. Vom 6. bis zum 9. Juli 2004 fand dazu eine Konferenz in Paris statt, auf der die Internationale Gesellschaft für phylogenetische Nomenklatur (International Society for Phylogenetic Nomenclature (ISPN)) begründet wurde, welche die in internationaler Zusammenarbeit erarbeiteten Regeln des „PhyloCode“ überwachen wird, der nach der Vorstellung seiner Verfasser langfristig alle bisherigen Nomenklatur-Regelwerke wie den botanischen (ICBN) oder zoologischen Nomenklatur-Code (ICZN) ablösen soll. Das Erstpublikationsdatum der Konferenzberichte wird als Startdatum des „PhyloCode “angegeben.
Ihren Befürwortern gilt die nach kladistischen Kriterien errichtete Taxonomie als einzige wirklich wissenschaftliche Methode zur Beschreibung, Einteilung und Benennung von Taxa, die im Gegensatz zur traditionellen Klassifikation, die als reines Schubladensystem angesehen wird, echte biologische Information widerspiegelt. Kritikern wie den oben erwähnten Ernst Mayr oder Gaylord Simpson gilt die kladistische Taxonomie dagegen als zu instabil und daher nicht praxistauglich.
Wie bereits zu Beginn angesprochen, muss die obenstehende Gegenüberstellung der beiden Taxonomien als idealisiert angesehen werden. Während es sich auch in der klassischen Taxonomie zunehmend durchsetzt, nach Möglichkeit monophyletische Taxa zu bilden und nur in Ausnahmefällen auf paraphyletische Gruppen auszuweichen, sind bei kladistisch arbeitenden Taxonomen zumindest auf Art- und Gattungsebene oft noch die klassischen binären Namen in Gebrauch. Auch bei höheren Taxa werden paraphyletische Gruppen zumindest informell noch weiterverwendet.
Operationale taxonomische Einheiten (MOTU)
Mit Hilfe molekularbiologischer Methoden können taxonomische Zugehörigkeiten erkannt werden. Nach DNA-Analyse und Vergleich in Genbibliotheken lassen sich molecular operational taxonomic units (MOTUs) definieren, was ermöglichen kann, Arten zu definieren. Damit konnten bei Saprolegnia 18 Arten bestätigt und weitere 11 potenzielle Arten identifiziert werden.
Literatur
- Gottlieb Wilhelm Bischoff: Wörterbuch der beschreibenden Botanik oder Die Kunstausdrücke, welche zum Verstehen der phytographischen Schriften nothwendig sind, lateinisch-deutsch und deutsch-lateinisch. 2. Auflage. Schweizerbart, Stuttgart 1857
- Willi Hennig: Grundzüge einer Theorie der Phylogenetischen Systematik. Deutscher Zentralverlag, Berlin 1950, Koeltz, Königstein 1980 (Repr.), ISBN 3-87429-188-X.
- Willy Hennig: Phylogenetic Systematics. Univ. of Illinois Press, London 1966, ISBN 0-252-06814-9.
- Lynn Margulis, Karlene V. Schwartz: Die fünf Reiche der Organismen. Spektrum d. Wiss., Heidelberg 1989, ISBN 3-89330-694-3.
- Lynn Margulis, Karlene V. Schwartz: Five Kingdoms, an Illustrated Guide to the Phyla of Life on Earth. Freeman, New York NY ³1998 (englisch. Orig.), ISBN 0-7167-3027-8.
- Ernst Mayr: Cladistic Analysis or Cladistic Classification. in: E. Mayr: Selected Essays. Evolution and the Diversity of Life. Harvard Univ. Press, Cambridge Mass 41997, ISBN 0-674-27105-X.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 16.02. 2026