Farbwahrnehmung

Die Farbwahrnehmung ist als Teilbereich des Sehens die Fähigkeit, Unterschiede in der spektralen Zusammensetzung des Lichts wahrzunehmen. Sie beruht darauf, dass das Auge über verschiedene Rezeptortypen verfügt, die jeweils für bestimmte Bereiche des Lichtspektrums empfindlich sind. Die Erregungsmuster dieser Rezeptorzellen bilden die Grundlage für die komplexe Weiterverarbeitung in Netzhaut und Gehirn, die schließlich zur Farbempfindung führt.
Dabei können unterschiedliche spektrale Zusammensetzungen des Farbreizes zur gleichen Farbwahrnehmung führen. Allein aus der wahrgenommenen Farbe kann daher nicht auf die Zusammensetzung des Farbreizes geschlossen werden. Ausschließlich Reize durch monochromatisches Licht einer bestimmten Wellenlänge sind auch charakterisierbar durch die wahrgenommene Farbe, als dessen Spektralfarbe.
Gegenstand des Artikels ist die naturwissenschaftliche Beschreibung der Farbwahrnehmung. Systeme der Ordnung von Farben werden in der Farbenlehre und die Messung von Farben in der Farbmetrik beschrieben.
Geschichte der Erforschung
- 1672: Isaac Newton entdeckt, dass Licht aus verschiedenen Farbanteilen zusammengesetzt ist, und beschreibt das Phänomen der Metamerie (unterschiedlich zusammengesetztes Licht kann denselben Farbeindruck hervorrufen).
- 1794: John Dalton berichtet über seine Farbsehschwäche. Etwas für Andere Rotes sah er als Verschattung, und er nahm Orange, Gelb und Grün nur als verschiedene Abstufungen von Gelb wahr. Die Rot-Grün-Blindheit wird daher auch als „Daltonismus“ bezeichnet. Dies ist nicht mit Rot-Grün-Sehschwäche zu verwechseln, wo Rot, Grün, Orange und Gelb unterscheidbar sind, aber nicht alle ihre Schattierungen.
- 1801: Thomas Young vermutet, dass die Möglichkeit, alle Farben aus drei Primärfarben zusammenzusetzen, auf physiologischen Vorgängen in der Netzhaut beruht, und schlägt drei Rezeptortypen vor, die zu den Primärfarben passen. Dieses Modell wurde um 1850 von Hermann von Helmholtz zur Dreifarbentheorie ausgebaut.
- 1855: James Clerk Maxwell identifiziert zwei Typen des „Daltonismus“ und erklärt sie mit Hilfe seiner Drei-Rezeptoren-Theorie.
- 1874: Der Physiologe Ewald Hering veröffentlicht seine Vierfarbentheorie als Gegenthese. Die Lösung des Widerspruchs und eine Weiterentwicklung der Sicht gelingt von Kries Ende des 19. Jahrhunderts mit der Kries-Zonentheorie.
Farbreiz, Farbvalenz und Farbempfindung


| Begriff | Wirkort | Wirkart | Fachgebiet |
|---|---|---|---|
| Farbreiz | Auge (Eintrittspupille) | elektromagnetische Strahlung | Entstehung von Farben / Optik |
| Farbvalenz | Zapfen und Stäbchen der Netzhaut | wellenlängenabhängige Empfindlichkeit der Rezeptoren der Netzhaut | Physiologie |
| Farbempfindung | Auge und Visueller Cortex | Farbwahrnehmung | Physiologie / Psychologie |
- Der Farbreiz ist die spektrale Verteilung jener Strahlungsleistung, für die die Zapfen der Netzhaut des Auges empfindlich sind. Dieser adäquate Reiz ist die physikalische Ursache von Farbvalenz und Farbempfindung. Der Farbreiz selbst ergibt sich aus den spektralen (und richtungsabhängigen) Reflexionscharakteristika des betrachteten Objektes, modifiziert durch die spektrale Zusammensetzung des einfallenden Lichtes.
- Die Farbvalenz ist die spektralspezifische physiologische Wirkung einer Strahlung. Sie ist charakterisiert durch die jeweiligen Erregungszustände der drei Zapfenarten des menschlichen Auges, die vom (physikalischen) Farbreiz abhängen.
- Die Farbempfindung entsteht durch ein Zusammenwirken der eintreffenden „mittleren“ Gesamthelligkeit und von abgleichenden Farbkonstanzleistungen des Gehirns. Die rezeptorbezogene trichromatische Reaktion – als unmittelbare Reizantwort der drei Zapfenarten – erreicht nicht das Bewusstsein. Entlang der Stationen der Signalverarbeitung von den Sinneszellen zum Großhirn werden die Parameterpaare Schwarz/Weiß (Hellwert), sowie Rot/Grün, Blau/Gelb (zwei konträre Buntpaare) geformt.
- Eine Spektralfarbe ist die Farbempfindung, die durch monochromatische Strahlung ausgelöst wird. Die durch Lichtbrechung an einem Prisma erzeugten Farben sind Spektralfarben.
Sichtbares Licht


Menschen können elektromagnetische Strahlung nur in dem Bereich visuell wahrnehmen, in dem das Sonnenspektrum den überwiegenden Anteil seiner Energie enthält. Der wahrnehmbare Bereich beginnt bei ca. 380 nm und endet bei etwa 780 nm. Da die Bezeichnung „Licht“ in der Physik gelegentlich auch für andere Abschnitte des elektromagnetischen Spektrums verwendet wird, hat sich für den sichtbaren Spektralbereich der Ausdruck „sichtbares Licht“ eingebürgert. Er umfasst die Farben von Blau-Violett über Grün und Gelb bis Dunkelrot. Sie können z. B. durch ein Prisma sichtbar gemacht werden. Einige Tiere können über den für Menschen sichtbaren Bereich hinaus sehen, beispielsweise Bienen, die auch das angrenzende nahe UV-Licht sehen. Unter optimalen Bedingungen und sehr hoher Lichtstärke können die Grenzen der menschlichen Wahrnehmung 310 nm (UV) bis 1100 nm (NIR) betragen.
In der Regel setzt sich Licht aus Lichtwellen unterschiedlicher Wellenlängen zusammen. Wenn alle Wellenlängen so vertreten sind, wie es näherungsweise für das Sonnenlicht gilt, spricht man von "weißem Licht". Hingegen wird Licht, das nur aus Strahlung einer bestimmten Wellenlänge besteht, monochromatisch genannt. Lichtquellen, die monochromatisches Licht aussenden, kommen in der Natur praktisch nicht vor. Der Regenbogen gilt nicht als Lichtquelle, da die Wassertropfen das Sonnenlicht reflektieren, wobei infolge der Lichtbrechung unter anderem auch monochromatische Farben entstehen. Technisch lassen sich monochromatische Lichtquellen jedoch realisieren, z. B. mit Natriumdampflampen oder Lasern. Der Mensch kann mit seinen Augen allerdings nicht unterscheiden, ob es sich bei Licht einer gewissen Farbe um ein monochromatisches Licht oder um ein (gleichwirkendes) „Gemisch“ von Licht unterschiedlicher Spektralbereiche handelt. Deswegen ist es wichtig, die physikalischen Eigenschaften des Lichts von der Farbempfindung begrifflich zu trennen.
Physiologie
Grundlagen
Das Wahrnehmungssystem muss mindestens zwei unterschiedliche Typen von „Lichtrezeptoren“ haben, um unterschiedliche Zusammensetzungen des Lichts feststellen zu können. Mit nur einer Art von Rezeptor sind Unterscheidungen nach der Wellenlänge nicht möglich, was W. Rushton 1970 als Prinzip der Univarianz herausstellte.
Der Mensch besitzt zwei verschiedene Systeme von visuellen Rezeptoren. Die Stäbchen sind zwar empfindlicher, aber es gibt davon nur einen Typ. Allein mit diesen Rezeptoren können folglich keine Farben unterschieden werden. Das zweite System besteht aus den Zapfen, den Rezeptoren des Farbsehens. Davon gibt es beim Menschen drei Typen mit unterschiedlicher spektraler Empfindlichkeit. Sie sind für das Tagessehen (Photopisches Sehen) verantwortlich. Ihre Reizantwort braucht eine Leuchtdichte von mindestens ca. 3 cd/m². Unterhalb dieser Schwelle sind ausschließlich Hell-Dunkel-Unterschiede durch die Stäbchen-Rezeptoren wahrnehmbar (skotopisches oder Nachtsehen). Für die Wahrnehmung in der Fovea centralis spielen allein Zapfen eine Rolle. Die extrafoveale Farbwahrnehmung kann unter bestimmten Dämmerungsbedingungen auch durch die Stäbchen beeinflusst werden, bei vollem Tageslicht sind diese jedoch durch die hohe Lichtintensität gesättigt und tragen nicht zur Farbwahrnehmung bei.
Sehpigmente
Möglich wird die Empfindlichkeit der Sinneszellen für verschiedene Wellenlängenbereiche durch die in ihrer Membran enthaltenen Moleküle spezifischer Sehpigmente. Diese bestehen aus einem Proteinanteil, Opsin genannt, und dem daran gebundenen Retinal-Molekül, das den lichtempfindlichen Anteil bildet. Letzteres ist als Ligand an Opsin kovalent gebunden. Trifft ein Photon mit passender Energie auf das Retinal-Molekül, so ändert es seine räumliche Struktur, und zwar vom gewinkelten 11-cis- zum gestreckten all-trans-Isomer des Moleküls. Diese Strukturveränderung wird als primäre photochemische Reaktion bezeichnet. Sie dauert etwa 2·10−14 sec und löst mehrere nachgeordnete Prozesse in der Sinneszelle aus, die das Signal erheblich verstärken und schließlich in einer Veränderung ihres Membranpotentials münden (Rezeptorpotential, hier durch Hyperpolarisation), welches dann eine nervliche Signalkette auslöst.
Die Sehpigmente der Stäbchen werden Rhodopsin genannt und bestehen aus Skotopsin und Retinal. Die Sehpigmente der drei Zapfenarten werden Iodopsine genannt. Sie bestehen jeweils aus einer der drei Arten von Photopsin und Retinal, die durch ihren leicht unterschiedlichen Aufbau unterschiedliche Reaktionsmaxima ergeben. Dies ist die Ursache für die unterschiedliche spektrale Empfindlichkeit der drei Typen von Zapfen (S-, M- und L-Typ).
Daneben finden sich beim Menschen auch photosensitive Ganglienzellen in der Netzhaut, die durch das Photopigment Melanopsin lichtempfindlich sind und ihre Signale an Neurone der Epiphyse und solche des Nucleus suprachiasmaticus im Hypothalamus weiterleiten, wo sie als Zeitgeber für die circadiane Rhythmik bestimmend werden.
Sinneszellen der Netzhaut
Photonen können also in den Sinneszellen der äußeren Schicht der Netzhaut, den Photorezeptoren, eine Verformung des Sehpigments bewirken und durch die anschließenden Prozesse Phototransduktion ein Rezeptorpotential auslösen. Dieses wiederum löst ein Signal aus für ein komplexes Netzwerk von Neuronen, das zwischen den Sinneszellen und den retinalen Ganglienzellen liegt, die die innerste Schicht der Netzhaut bilden. Nach massiver Querverarbeitung in diesem Netzwerk gehen Signale dann an die Ganglienzellen und werden von dort aus über die Neuriten des Sehnervs an das Gehirn weitergeleitet. Hier schließlich tragen sie, nach weiteren Prozessen, zur Wahrnehmung der erlebten Farbeindrücke bei.
Für die visuelle Wahrnehmung beim Menschen sind dabei, wie schon erwähnt, zwei Systeme von Photorezeptoren zu unterscheiden:
- Die Stäbchen sind noch bei geringer Lichtintensität unter 0,1 cd/cm² aktiv und für das Nachtsehen verantwortlich.
- Die drei verschiedenen Arten von Zapfen
registrieren unterschiedliche Farbvalenzen.
Jede Zapfenart hat einen spezifischen spektralen Empfindlichkeitsbereich.
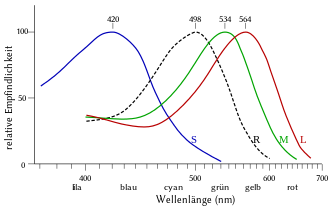
- S-Zapfen (S für Short) sind empfindlich für kürzere Wellenlängen (etwa 400–500 nm). Das Absorptionsmaximum liegt bei etwa 420 nm, was einem violettgetönten Blau entspricht. S-Zapfen werden auch Blau-Zapfen genannt und sind beim Menschen nur mit einem Anteil von zwölf Prozent aller Zapfen vertreten.
- M-Zapfen (M für Medium) sind empfindlich für mittlere Wellenlängen (etwa 450–630 nm). Das Absorptionsmaximum liegt bei etwa 530 nm, was einem türkisgetönten Grün entspricht. M-Zapfen werden auch Grün-Zapfen genannt.
- L-Zapfen (L für Long) sind empfindlich für längere Wellenlängen (etwa 500–700 nm). Das Absorptionsmaximum liegt etwa bei 560 nm, was einem grünlichen Gelb entspricht. L-Zapfen werden auch Rot-Zapfen genannt.
Die unterschiedlichen Absorptionsspektren kommen dadurch zustande, dass die drei Zapfentypen jeweils eine andere Unterform des Sehpigments enthalten. Deren Proteinanteil – das so genannte Opsin – wird von Genen codiert, die sich auf dem 7. Chromosom (im Falle von S-Opsin) und auf dem X-Chromosom (im Falle von L- und M-Opsin) befinden. Da für die Rot-Grün-Schwäche ein mutiertes L- oder M-Opsin-Gen verantwortlich ist, führt deren gonosomale Vererbung dazu, dass die Rot-Grün-Schwäche bei Männern sehr viel häufiger ist als bei Frauen.
Die Absorptionskurven aller drei Zapfenarten sind verhältnismäßig breit und überlappen stark. Allein ein Zapfentyp liefert keine hinreichende Information für Farbensehen. Dies wird erst möglich durch die neuronale Weiterverarbeitung der Erregungsmuster von mindestens zwei verschiedenen Zapfentypen.
Die Zapfendichte ist ungefähr in der Netzhautmitte, am Punkt des schärfsten Sehens (Fovea centralis), am größten. Die Dichte nimmt nach außen hin ab, und am Rand des Gesichtsfeldes sind kaum noch Zapfen vorhanden, dafür viele Stäbchen. In der Fovea centralis gibt es wiederum keine Stäbchen. Beispielsweise kann man schwach leuchtende Sterne nachts nur sehen, wenn man etwas an ihnen „vorbeischaut“, und dann ohne Farbe.
Neuronale Verarbeitung von Farbreizen

links:
▪ RPE retinales Pigmentepithel
▪ OS Außensegmente der Photorezeptorzellen
▪ IS Innensegmente der Photorezeptorzellen
▪ ONL äußere nukleäre Schicht
▪ OPL äußere plexiforme Schicht
▪ INL innere nukleäre Schicht
▪ IPL innere plexiforme Schicht
▪ GC Ganglienzellschicht
rechts:
▪ BM Bruch-Membran, P Pigmentepithelzelle
▪ R Stäbchen, C Zapfen
▪ ◁ Membrana limitans externa
▪ H Horizontalzelle, Bi Bipolarzelle
▪ M Müllerzelle, A Amakrinzelle
▪ G Ganglienzelle, Ax Axone
Bereits in der Retina, die aus dem Augenbecher entstanden embryologisch ein Teil des Hirns ist, finden erste Schritte der Informationsverarbeitung statt. Je eine Gruppe von Sinneszellen (Zapfen oder Stäbchen) konvergiert über zwischengeschaltete Nervenzellen (Bipolare Zellen, Horizontalzellen, und Amakrinzellen) auf jeweils eine retinale Ganglienzelle, das 3. Neuron. Eine solche Gruppe von Sinneszellen bildet ein rezeptives Feld, und man unterscheidet darin ein Zentrum und dessen Peripherie. Die Photorezeptoren des Zentrums wirken gegensinnig zu denen der Peripherie auf die nachgeschaltete Ganglienzelle. Ist das Zentrum erregend und die Peripherie hemmend, so spricht man von einer On-Zentrum-Ganglienzelle, im umgekehrten Fall von einer Off-Zentrum-Ganglienzelle. Diese Art der Verschaltung dient der Kontrastverstärkung.
Im Wesentlichen lassen sich drei Teilsysteme unterscheiden.
- Diffuse Bipolarzellen geben sowohl Signale von L- als auch von M-Zapfen auf sogenannte parasol-Ganglienzellen (auch M-Zellen genannt) weiter, deren Axone in die magnozellulären Schichten des Corpus geniculatum laterale (CGL) ziehen. Sie zeigen eine breite spektrale Ansprechbarkeit. Die Information, die sie weiterleiten, ist also achromatisch und dient vermutlich vor allem der Unterscheidung von Hell und Dunkel.
- Die sogenannten midget-Ganglienzellen (auch P-Zellen genannt) empfangen hingegen Signale (über midget-Bipolarzellen) von lediglich einem L-Zapfen oder einem M-Zapfen im Zentrum. Die rezeptiven Felder dieser Zellen sind sehr klein und reagieren unterschiedlich auf langwelliges und mittelwelliges Licht. Die Axone der midget-Ganglienzellen ziehen in die parvozellulären Schichten des CGL. Sie verarbeiten vor allem den Rot/Grün-Kontrast. Evolutionsgeschichtlich handelt es sich hierbei um das jüngste Teilsystem, erst bei den Primaten sind die Opsine der L- und M-Zapfen durch eine Genduplikation entstanden.
- Auf die bistratified-Ganglienzellen konvergieren blue-Bipolarzellen und bilden ein On-Zentrum von S-Zapfen aus, diffuse Bipolarzellen leiten Signale von L- und M-Zapfen, die hemmend wirken (off). Damit können vor allem Blau/Gelb-Kontraste hervorgehoben werden. Die Axone dieser retinalen Ganglienzellen projizieren auf die koniozellulären (Unter-)Schichten des CGL.
In allen drei Fällen wirken Horizontalzellen bei der Ausbildung der rezeptiven Felder mit und Amakrinzellen modulieren den Signalfluss zu den Ganglienzellen.
Neben der Differenzierung der Farbqualitäten sind weitere Aufarbeitungsprozesse bekannt.
- Das schnell arbeitende Rot-Grün-System der stammesgeschichtlich gemeinsam entwickelten M- und L-Zapfen dient auch der Hervorhebung von Kanten im Bildmuster. Dabei wird die Differenz von L-(Rot) und M-(Grün)Signalen mit der Summe beider verglichen. Wenn unter Laborbedingungen beide Zapfentypen mit rotem und grünem Licht gleicher Stärke gereizt werden (Isoluminanz), nimmt das Wahrnehmungsvermögen für scharfe Kanten stark ab (minimally distinct border-Phänomen).
- Das weniger rasch arbeitende Blau-Gelb-System ist darüber hinaus noch für die Farbkonstanz zuständig.
- Das Signal des Rotzapfens alleine wird vermutlich zur Bewegungsdetektion insbesondere bei langsamen Abläufen verwendet.
Spezielle Besonderheiten der Farbwahrnehmung
Metamere Farbgleichheit
Farbreize werden durch Kombinationen verschiedener Wellenlängen des elektromagnetischen Spektrums erzeugt. Derselbe Farbreiz kann durch unterschiedliche Kombinationen erzeugt werden. Dieser Effekt wird Metamerie genannt. Zwei Farbproben können daher (unter gleicher Beleuchtung) völlig identisch aussehen, obwohl sie unterschiedlich spektrale Anteile des Lichtes absorbieren. Wenn man die Farbproben mit farbigem Licht beleuchtet – also mit Licht, in dem Spektralanteile fehlen – kann der Unterschied sichtbar werden, sofern der fehlende Spektralanteil in der einen Farbprobe mehr zu ihrem Aussehen beiträgt als in der anderen. Dies ist ein Problem bei der Herstellung von Dingen und Gegenständen aus unterschiedlichen Materialien, welche auch unter unterschiedlichen Beleuchtungsverhältnissen gleichfarbig aussehen sollen.
Farbkonstanz
Die Farbkonstanz zählt zur Gruppe der sogenannten Konstanzphänomene der menschlichen Wahrnehmung, neben u. a. der Formkonstanz und der Größenkonstanz. Als Farbkonstanz bezeichnet man die Eigenschaft des Sehsinns, die Körperfarbe von Gegenständen als nahezu unabhängig von Veränderungen der Farbspektra natürlicher Beleuchtung wahrzunehmen. Derartige Veränderungen geschehen in Zeitabständen von Jahreszeiten, Tageszeiten, Bewölkungsänderungen, und Schattenwurf bei Orts- oder Blickwechsel.
Farbkonstanz wurde auch bei Fischen und Bienen nachgewiesen. Hier wird der Vorteil der Fähigkeit besonders anschaulich, da die Farbe der Beleuchtung unter Wasser und im Bereich von Blüten schnell und intensiv wechseln kann. Die Nahrungssuche wird hier wesentlich vereinfacht, oder überhaupt erst ermöglicht, wenn das Gesuchte immer in nahezu denselben Farben gesehen wird.
In der Fotografie kann man Effekte von Wechsel in natürlicher, oder auch künstlicher, Beleuchtung durch Aufnahmen mit Kunstlichtfilmen bei Tag oder Tageslichtfilmen bei Kunstlicht nachstellen. Mit einer Digitalkamera lassen sich solche Effekte bei Veränderungen des Weißabgleichs beobachten.
Neurophysiologie der Farbkonstanz
Die Aufklärung des Mechanismus der Farbkonstanz gilt als besondere Herausforderung innerhalb der Neurowissenschaft des Sehsinnes. Da die Fähigkeit darauf gründet, dass neuronale Repräsentationen von weit auseinander liegenden Bereichen des Gesichtsfeldes sich gegenseitig beeinflussen, müssen höhere kortikale Prozesse beteiligt sein. Eine nachweisliche Schlüsselrolle für den Mechanismus der Farbkonstanz spielt das visuelle Cortexareal V4. Doch sind auch Anpassungsprozesse auf tieferen Ebenen beteiligt, so schon innerhalb der Netzhaut. Darüber hinaus ist bekannt, dass es beim Menschen interindividuelle Unterschiede in der Realisierung der Farbkonstanz gibt. Die Sichtweisen von Malern, beispielsweise impressionistischer wie Claude Monet bei seiner berühmten Bilder-Reihe mit Ansichten der Kathedrale von Rouen, verdeutlichen, dass hierbei sowohl Aufmerksamkeits- als auch Lernprozesse eine Rolle spielen.
UV-Wahrnehmung beim Menschen
Das Rhodopsin der menschlichen Stäbchen hat zwei Absorptionsmaxima, eines im sichtbaren Bereich bei 500 nm (türkis) und ein Nebenmaximum im UV-Bereich bei 350 nm. Durch die Absorption von UV-Licht in der Augenlinse wird beim menschlichen Auge normalerweise ein Reiz im UV-Bereich weitgehend verhindert. Für die Netzhaut, die durch die energiereiche UV-Strahlung geschädigt werden kann, stellt dies eine Schutzfunktion dar. Menschen, denen die Linse entfernt wurde (z.B. wegen Grauen Stars) können UV-Lichtreize als hell wahrnehmen, ohne dies jedoch als Farbe zu sehen.
Defekte der Farbwahrnehmung
Farbfehlsichtigkeit tritt in verschiedenen Formen auf.
- Rotblinde ohne Rotrezeptoren werden als Protanope (gr. protos, erster; gr. an-, nicht; gr. ope Blick) bezeichnet
- Grünblinde als Deuteranope (gr. deuteros, zweiter), sie weisen beide das Phänomen der Dichromasie auf, besitzen also nur zwei statt drei Zapfentypen.
- Rotschwäche (Protanomalie) und Grünschwäche (Deuteranomalie) beruhen auf veränderten Empfindlichkeiten der entsprechenden Rezeptoren.
Diese Fehlsichtigkeiten treten bei Veränderungen der Opsin-Gene auf. Aber auch Linsenverfärbungen (Vergilbungen) können die Farbwahrnehmung beeinträchtigen. Es gibt unterschiedliche Möglichkeiten Defekte zu erkennen, wie Ishihara-Farbtafel oder der Farnsworth-Test. Ob jemand für einen bestimmten Beruf (Pilot) geeignet ist, kann außerdem mit anderen speziellen Tests (Beyne-Lanterntest) ermittelt werden.
Farbwahrnehmung im Tierreich

„Farbig“ sehen
Farbensehen unterscheidet sich beträchtlich zwischen den Tierarten. In der Evolutionsgeschichte hat sich das Sehen mehrfach – und unabhängig voneinander – entwickelt. Es gibt Unterschiede in der Anzahl der Rezeptortypen und in deren spektraler Empfindlichkeit. Die meisten Säugetiere haben zwei verschiedene Typen von Farbrezeptoren, einige Primaten drei, Reptilien und die in der Evolution aus diesen heraus entwickelten Vögel oft vier.
Verbreitung im Tierreich
Wirbellose
Gliederfüßer
- Bei Insekten wurde das Farbensehen insbesondere bei der Honigbiene untersucht. Karl von Frisch hat gezeigt, dass man Bienen nach ihren Farbempfindungen „fragen“ kann, indem man sie auf farbige Plättchen mit Futterbelohnung dressiert. Für den Nachweis echten Farbensehens reicht es dabei nicht aus, dass ein Tier immer wieder auf die einmal als futterträchtig erfahrene Farbe zurückkehrt, denn es könnte ja die Graustufe gelernt haben. Der Sinnesreiz Farbe wird nur dann erkannt, wenn sie unabhängig von der Helligkeit immer wieder gewählt wird. Frisch prüfte dies, indem er den Bienen Farbplättchen verschiedener Helligkeit der belohnten Farbe in Konkurrenz mit anderen Farben zur Auswahl anbot, und feststellte, dass die „Farbe“ bei der Entscheidung Priorität hat.
- Der Fangschreckenkrebs Neogondodactylus oerstedii besitzt acht verschiedene Rezeptortypen im sichtbaren und vier im Ultraviolett-Bereich.
Wirbeltiere
Niedere Wirbeltiere
Niedere Wirbeltiere, und unter den Säugetieren die Beuteltiere, verfügen meist über vier Zapfentypen. Sie werden daher Tetrachromaten genannt. Neben den L-, M- und S-Zapfen verfügen sie über einen Ultraviolett-Zapfen, der im Bereich von weniger als 380 nm reagiert. Da man dieses – im Vergleich zum Menschen komplexere – tetrachromatische Farbsystem bei Beuteltieren, Vögeln und Fischen findet, geht man davon aus, dass es einen evolutionär frühen Typus innerhalb der Wirbeltier-Farbsysteme darstellt.
Knochenfische
In Anpassung an die unterschiedlichen Beleuchtungsverhältnisse ihrer Lebensräume haben die verschiedenen Arten der Knochenfische unterschiedliche Systeme ausgebildet. Die meisten bisher darauf untersuchten Fische sind Tetrachromaten. Die Zahl der Zapfen und deren Absorptionsmaxima passen in der Regel zu ihrer Lebensweise: Mit zunehmender Tiefe in Gewässern ist auf Grund der stärkeren Absorption von lang- und kurzwelligem Licht die Beleuchtung zunehmend einfarbiger (monochromatisch). In klaren Meeren oder Seen erreicht der blaue Anteil des Lichtes Tiefen von über 60 Metern. In Süßwasserseen mit einer hohen Planktondichte herrscht in Tiefen von 25 Metern gelbgrünes Licht vor. In Schwarzwasserflüssen und Moorseen erreicht der Rotanteil des Lichtes höchstens eine Tiefe von drei Metern. Gleichzeitig nimmt bei allen Gewässern die Intensität des Lichtes mit der Tiefe ab. So besitzen dämmerungsaktive oder in dunklen Regionen lebende Fische vorwiegend im Rotbereich empfindliche Zapfen, während tagaktive, in den oberen, lichtdurchfluteten Regionen lebende Fische mehr Blau- und Grünzapfen aufweisen.
- Stäbchenmonochromaten besitzen keine Zapfen. Sie können bei sehr geringen Lichtintensitäten sehen, dafür aber nur Graustufen. Das hellste Grau liefern Objekte in Grüntönen.
- Dichromaten haben zusätzlich zwei verschiedene Zapfentypen. Beispiel: Gemeine Goldmakrele (Coryphaena hippurus).
- Trichromaten besitzen ähnlich wie der Mensch drei Zapfentypen. Beispiel: Buntbarsch (Cichlasoma longinasus).
Ob Di- und Trichromaten auch unterschiedliche Farben wahrnehmen und unterscheiden können, hängt von der weiteren neuronalen Verarbeitung in Netzhaut und Gehirn ab.
Vögel
Hühner besitzen neben dem Rhodopsin der Stäbchen vier Zapfenpigmente: Rot (Empfindlichkeitsmaximum bei ca. 570 nm), Grün (ca. 510 nm), Blau (ca. 450 nm) und Violett (ca. 420 nm). Zusätzlich befindet sich im Pinealorgan (Zirbeldrüse/Epiphyse) ein weiteres Pigment, das Pinopsin (ca. 460 nm).
Vögel und ebenso Reptilien haben in ihren Zapfen mit Carotinoiden gefärbte und farblose Öltröpfchen, die als Farbfilter wirken. Diese Filter engen die Absorptionsspektren der Zapfen ein und verbessern damit die Unterscheidbarkeit von Farben. Säugetiere, und somit auch der Mensch, besitzen diese Farbfilter nicht.
Säuger
- Mäuse haben neben dem Stäbchenpigment Rhodopsin nur zwei Zapfenpigmente für Grün (Absorptionsmaximum ca. 510 nm) und Ultraviolett (ca. 350 nm). Die Zapfenpigmente sind zudem ungleich verteit. In der oberen Hälfte der Netzhaut, die den Boden betrachtet, finden sich neben Stäbchen nur Grün-Zapfen, in der unteren Hälfte der Netzhaut, die den Himmel betrachtet, neben Stäbchen nur UV-Zapfen. Dies ist eine Anpassung zur Nahrungssuche bzw. zum leichteren Erkennen der UV-Muster in den Gefiedern von Beutegreifern.
- Dass Hunde keinen Farbsinn haben, sie also Schwarz-Weiß sehen, wird selbst in populärwissenschaftlichen Beiträgen noch immer behauptet. Doch auch der Haushund verfügt über zwei Zapfenarten mit Empfindlichkeiten im grünen und blauen Spektralbereich.
- Primaten können „farbig“ sehen. Wie Untersuchungen an Affen am japanischen Nationalen Forschungsinstitut in Tsukuba ergaben ist die Fähigkeit, Farbtöne unabhängig von der Helligkeit wahrzunehmen, nicht angeboren. Dies stellte man bei Affen fest, die in monochromatischem Licht aufgewachsen waren. Sie konnten ein farbiges Objekt immer dann nicht wiedererkennen, wenn dies bei abweichenden Beleuchtungsverhältnissen Licht unterschiedlicher Wellenlänge reflektierte.
Evolution der Zapfentypen bei Landwirbeltieren

| Zapfentypen | UV | S | M | L | Stäbchen |
|---|---|---|---|---|---|
| Mensch | - | 424 nm | 530 nm | 560 nm | k. A. |
| Mensch | - | 420 nm | 535 nm | 565 nm | k. A. |
| Mensch | - | 420 nm | 530 nm | 560 nm | 500 nm |
| Rhesusaffe | - | * | 540 nm | 565 nm | 505 nm |
| Pferd | - | 428 nm | 539 nm | - | - |
| Vögel | 370 nm | 445 nm | 508 nm | 565 nm | - |
| Goldfisch | 356 nm | 447 nm | 537 nm | 623 nm | - |
Anmerkung
- Absorption wird hier als Anzahl der von einem Zapfen pro Sekunde aufgenommenen Photonen bestimmt.
- Die genannten Absorptionsmaxima sind nur Richtwerte; Unterschiede gibt es nicht nur zwischen den Arten, sondern auch von Individuum zu Individuum.
Vögel besitzen vier Zapfentypen, deren Absorptionsmaxima bei 370 nm (UV-Typ), 445 nm (S-Typ), 508 nm (M-Typ) und 565 nm (L-Typ) liegen. Auf Grund von Vergleichen der DNA-Sequenzen verschiedener Opsin-Typen bei verschiedenen rezenten Tieren nimmt man an, dass auch die gemeinsamen Vorfahren der Vögel und Säuger vier Zapfentypen besaßen. In einer frühen Phase der Säugerevolution gingen die mittleren S- und M-Typen verloren. Es wird angenommen, dass diese Tiere nachtaktiv waren und deswegen diese Veränderung im Sehsystem tolerieren konnten. Wie eine Abschätzung der Evolutionsdauer mit Hilfe der molekularen Uhr ergab, entstand mit dem Übergang zur Tagaktivität vor 30 bis 40 Mio. Jahren bei den Vorfahren der Primaten der Alten Welt durch Genduplikation ein dritter Zapfentyp, so dass wieder ein M-Typ (530 nm) zur Verfügung stand, dessen Absorptionsmaximum sich aber nur wenig vom L-Typ (560 nm) unterscheidet. Ein selektiver Vorteil bestand möglicherweise darin, dass sich mit drei Zapfentypen Früchte als Nahrungsquelle besser unterscheiden lassen als mit zwei.
Ultraviolett-Wahrnehmung
Viele Insekten, Vögel, Eidechsen, Schildkröten und Fische haben Rezeptoren in ihrer Netzhaut, die auch bei Licht mit Wellenlängen kürzer als 400 nm – also durch Ultraviolett – gereizt werden. Aber auch manche Wirbellose verfügen über Rezeptoren für UV: Sir John Lubbock, ein Freund und Nachbar von Charles Darwin, stellte schon vor 1882 fest, dass Ameisen unter Ultraviolett (UV) ihre Puppen aufnehmen und aus dem Strahlungsbereich tragen. Karl von Frisch erkannte in den 1950er Jahren, dass Bienen und Ameisen UV-Licht als Farbe wahrnehmen.

W: Weißpunkt, Wellenlängen in nm
Auf Grund des vierten Zapfentyps, der im Ultraviolett (UV) oder Violett (V) sein Absorptionsmaximum hat, können bestimmte Tiere wie einige Insekten, nahezu alle Fische (Goldfisch), Reptilien, die Ursäuger Australiens und Vögel mehr Farben unterscheiden als der Mensch. Man spricht von Tetrachromasie. Untersuchungen am Wellensittich (Melapsittacus undulatus) ergaben, dass der Vogel nicht nur die Farben, die auch der Mensch unterscheidet, wahrnehmen kann, sondern darüber hinaus auch Mischungen mit unterschiedlichem UV-Anteil. So unterscheidet ein Vogel je nach UV-Anteil zum Beispiel bei einem bestimmten Blau verschiedene Farben, wo der Mensch nur eine einzige wahrnehmen kann.
Aus der Anzahl der Zapfentypen kann jedoch keineswegs geschlossen werden, welche Fähigkeiten der Farbunterscheidung ein Tier hat. Dies hängt sehr stark von der Weiterverarbeitung der Zapfenreaktionen in Netzhaut und Gehirn ab und kann erst durch Verhaltensexperimente untersucht werden.
Bedeutung
Die Fähigkeit, Ultraviolett wahrzunehmen, spielt für einige Vögel bei der Balz eine Rolle.
- Messungen der UV-Reflexion ergaben, dass von 139 Arten, bei denen sich Männchen und Weibchen für das menschliche Auge nicht unterscheiden lassen, sich bei mehr als 90 % der Arten die Geschlechter im UV-Muster unterscheiden.
- Bei Männchen 108 australischer Vogelarten reflektieren diejenigen Stellen des Gefieders, die bei der Balz eine Rolle spielen, mehr UV als andere Gefiederareale.
- Bei der Blaumeise (Parus caeruleus) wählen die Weibchen bevorzugt diejenigen Männchen, die am stärksten im Ultraviolett-Bereich leuchten. Da die UV-Reflexion von der Mikrostruktur der Federn abhängt, kann sie Auskunft über die Gesundheit der Männchen geben.
- Beim Azurbischof (Guiraca caerulea) besetzen die Männchen mit der höchsten UV-Reflexion die größten und ertragreichsten Reviere und füttern ihre Jungen am häufigsten.
Aber auch beim Nahrungserwerb spielt die Wahrnehmung von Ultraviolett oder dessen jeweilige Ausprägung eine Rolle.
- Die Oberfläche von vielen Früchten reflektiert Ultraviolett. Dadurch ist es für Tiere mit UV-Wahrnehmung leichter, solche Früchte zu finden oder ihre jeweilige Qualität abzuschätzen.
- Turmfalken (Falco tinnunculus) entdecken die Spur ihrer Beute (z. B. von Erdmäusen) anhand deren Markierungen, da Urin und Kot Ultraviolett reflektieren.
Reife gelbe Bananen fluoreszieren in ultraviolettem Licht blau, im Gegensatz zu den Blättern dieser oder anderer Pflanzen. Das ist ein Hinweis darauf, dass manche Bananen fressenden Tiere diese Eigenschaft mit Hilfe ihrer UV-Wahrnehmung nutzen könnten, um die Reife der Früchte abzuschätzen.
.jpg)
Kulturelle Einflüsse
Zwar sind die neuronalen Wege und Mechanismen der Verarbeitung von Farbinformationen beim Menschen im Prinzip bekannt, die Entstehung des Bewusstseins über Farbe ist jedoch – ähnlich wie die Entstehung des Bewusstseins über andere Dinge – von vielen weiteren, und oft unbekannten, Einflüssen abhängig. „Offenbar ist es nicht die primäre entwicklungsgeschichtliche Aufgabe des Sehsinnes, ästhetische Empfindungen hervorzubringen. Vielmehr scheint es seine wichtigste Aufgabe zu sein, dem Individuum durch sichere Orientierung und durch optimales Erkennen das Überleben zu gewährleisten. Deshalb hat sich der Sehsinn so herausgebildet, dass er sich an die Beleuchtungsumstände weitestgehend anpassen kann.“ (Harald Küppers) Hinweise, dass auf dieser Stufe der Wahrnehmung kulturelle und damit durch Lernprozesse beeinflusste Unterschiede bestehen, gibt die Benennung von Farben und die Einteilung des Farbspektrums in Farbgruppen.
Farbnamen und Farbsortierungen
- Empedokles fasst Weiß und Schwarz als Farben auf.
- Aristoteles setzt in seinem Werk De sensu („Über die Sinne“) die Helligkeit der Luft der Farbe Weiß von Körpern gleich, Dunkelheit entspricht der Farbe Schwarz. So kommt er zu der Annahme, dass Farben sich aus unterschiedlichen Mischungen von Weiß und Schwarz zusammensetzen.
Nach diesen Vorstellungen wurden die Farben bis ins 17. Jahrhundert nach einer Helligkeitsskala angeordnet: Weiß – Gelb – Rot – Blau – Schwarz. Während heutzutage eine Farbe durch Farbton, Sättigung und Helligkeit definiert ist, sah man den Farbton bis dahin nur als Folge von Helligkeit an. Goethe wurde noch von dieser Auffassung beeinflusst, als er Newtons Ergebnisse ablehnte und seine eigene Farbenlehre sogar noch höher als sein literarisches Schaffen einschätzte.
Die Sichtweise der Sortierung nach Helligkeit findet teilweise eine Entsprechung in der Etymologie des Begriffs Gelb, der auf eine indogermanische Wurzel mit der Bedeutung ›hell, glänzend‹ zurückgeht.
Farbkategorien
In sogenannten natürlichen Sprachen wird die riesige Anzahl unterscheidbarer Farbnuancen einigen wenigen Farbkategorien zugeordnet, zum Beispiel in europäischen Sprachen oft: Violett, Blau, Grün, Gelb, Orange, Rot, Rosa, Braun. Untersuchungen ergaben, dass die Berimos auf Papua-Neuguinea nur fünf Kategorien verwendeten. So ordneten sie einen weiten Bereich an Farbnuancen, der von Europäern in die beiden Kategorien Grün und Blau eingeteilt wird, nur einem Begriff zu.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 18.03. 2026