Stäbchen (Auge)
Stäbchen oder Stäbchenzelle, griechisch-lateinisch Neuron bacilliferum, heißt eine Art von Fotorezeptoren in der Netzhaut des Wirbeltierauges mit stabförmigem Fortsatz, dem Stäbchen, anatomisch Bacillum retinae (lateinisch bacillum ‚Stäbchen‘). Stäbchenzellen sind Neuronen, die als spezialisierte Sinneszellen dem skotopischen Sehen bei geringer Helligkeit, Nachtsehen oder Dämmerungssehen dienen. Mit diesen sehr empfindlichen Lichtsinneszellen können auch schwache Lichtreize aus der Außenwelt in ein für das Gehirn verwertbares Signal umgewandelt werden. Die Auswertung allein der Signale von Rezeptorzellen mit Stäbchen ermöglicht ein Hell-Dunkel-Sehen; da die Rezeptoren dieses einen Typs alle auf Licht desselben bestimmten Wellenlängenbereichs reagieren, spricht man hier auch von monochromatischem Sehen. Viele Tiere besitzen zusätzlich die analog aufgebauten, weniger empfindlichen Zapfen, von denen es verschiedene Typen gibt, die für eine Farbwahrnehmung, das sogenannte Photopische Sehen, notwendig sind.
Aufbau und Funktion
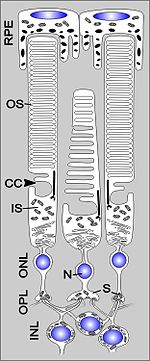
Im Aufbau sind Stäbchen und Zapfen ähnlich organisiert und bestehen aus einem Zellkörper, einer Synapse sowie einer Zellspezialisierung: dem Innen- und Außensegment. Im Außensegment („Outer segment“, OS) findet die visuelle Signaltransduktion durch die Sehfarbstoffmoleküle statt. Diese setzen sich aus einer chromophoren Gruppe (Retinal) und einem Glykoprotein (Opsin) zusammen. Diese Moleküle sind in vielen (> 1000) membranösen Scheiben („Disks“) eingelagert. Die Außensegmente der Stäbchen sind lang, schmal und grenzen an das retinale Pigmentepithel (RPE), welches abgeschnürte, alte Membranstapel phagocytiert. Ein Außensegment ist über ein modifiziertes Cilium in dezentraler Lage, das Verbindungscilium („Connecting cilium“, CC), mit dem Innensegment verbunden. Neun Mikrotubuli-Dupletts in nonagonaler Anordnung bilden die innere Struktur dieses unbeweglichen Ciliums. An dieses schließt sich das stoffwechselaktive Innensegment („Inner segment“, IS) an, welches noch in das an Mitochondrien reiche Ellipsoid und in das Myoid mit dem Endoplasmatischen Retikulum (ER) unterteilt ist. Hier erfolgt unter anderem die Proteinbiosynthese. Die folgende Schicht ist die äußere Körnerschicht („Outer nuclear layer“, ONL), welche den Zellkörper mit dem Zellkern (Nucleus, N) beinhalten. Von diesem geht ein Axon aus, welches mit einer Synapse (S) in der äußeren plexiformen Schicht („Outer plexiform layer“, OPL) endet. Die Synapsen der Photorezeptoren sind sogenannte „Ribbon-Synapsen“, band- oder plattenartige Strukturen direkt an der aktiven Zone der Präsynapse.
An die Ribbon-Struktur sind viele synaptische Vesikel gekoppelt, und es können im Vergleich zu normalen Synapsen eine weit höhere Anzahl von Vesikel pro Zeiteinheit ausgeschüttet werden. Im Dunkeln erfolgt eine fortwährende Ausschüttung des Neurotransmitters Glutamat. Dieser wirkt in der Regel exzitatorisch auf die Postsynapsen von Horizontal- und Bipolarzellen. Trifft Licht auf die Photorezeptorzelle, werden Ionenkanäle in der Zellmembran geschlossen, ausgelöst durch die Signaltransduktionskaskade. Die Photorezeptorzelle hyperpolarisiert und schüttet den Neurotransmitter nicht weiter aus. In der Folge werden die Ionenkanäle der nachgeschalteten Zellen geöffnet und so der Impuls an diese übertragen.
Lichtempfindlichkeit
.svg.png)
Die Stäbchen des Menschen enthalten eine Form des Sehpigments Rhodopsin, die am empfindlichsten für Licht mit einer Wellenlänge von etwa 500 nm (blaugrün) ist. Diese Sinneszellen sind hauptsächlich für das Sehen in der Dämmerung und bei Nacht wichtig, da sie schon bei geringer Lichtintensität arbeiten. Durch die Stäbchen können keine Farben unterschieden werden, weil im Gegensatz zu den Zapfen alle Stäbchen dasselbe Empfindlichkeitsspektrum aufweisen. Im Außenbereich der Netzhautmitte (5–6 mm) überwiegt die Zahl der Stäbchen, wodurch der Mensch bei Dämmerung in der Peripherie besser sieht als im Zentrum. Insgesamt befinden sich im menschlichen Auge etwa 120 Millionen Stäbchen und etwa 6 Millionen Zapfen.
Die größere Lichtempfindlichkeit der Stäbchen gegenüber den Zapfen hat hauptsächlich zwei Ursachen:
- Zum einen sind die lichtempfindlichen Pigmentscheiben im oberen Teil der Stäbchen lichtempfindlicher. Bereits ein einzelnes absorbiertes Photon führt nach einer Reihe von intrazellulären Prozessen zu einer Membranspannungsänderung von etwa 1 mV. Zapfen benötigen hingegen eine wesentlich größere Photonenanzahl (mindestens etwa 200), um eine Erregung an die nachgeschalteten Zellen weiterzuleiten.
- Der zweite Grund liegt in der neuronalen Verschaltung der Rezeptoren mit nachgeschalteten Zellen. Grob gesagt leiten viele Stäbchen ihr Signal an eine einzige Ganglionzelle (über Bipolarzellen etc.) weiter, während ein Zapfen in vielen Fällen auch nur auf jeweils eine Ganglionzelle ableitet. Das heißt die Information der Stäbchen konvergiert ungleich stärker als jene der Zapfen. Darin liegt auch der Grund für die schlechte räumliche Auflösung des Stäbchensehens (beispielsweise in der Nacht). Erhält eine Ganglienzelle (über welche die Information letztendlich weiter Richtung Gehirn geleitet wird) ein Stäbchensignal, so kann dieses von vielen verschiedenen Stäbchen, die mit ihr Synapsen bilden, stammen, und der Punkt auf der Netzhaut, wo das Bild abgebildet wird, ist somit relativ vage. Erhält eine Ganglionzelle hingegen Zapfeninformation, so kann der Lichtpunkt sehr gut auf der Retina lokalisiert werden, da nur sehr wenige Zapfen mit ihr verbunden sind.
Erregungsweiterleitung bei Stäbchen und Zapfen
Die übergroße Mehrheit der Nervenzellen (Neurone) leitet ihre Information über sogenannte Aktionspotentiale an andere Neurone weiter. Vereinfacht gesprochen wird durch die Reizung eines Neurons in ihm eine Spannungsänderung bewirkt (die eigentlich negativ geladene Zelle wird für kurze Zeit positiv geladen), was dazu führt, dass das Neuron über eine synaptische Verbindung Botenstoffe (Neurotransmitter) ausschüttet. Diese Neurotransmitter binden an Rezeptoren des nachgeschalteten Neurons und führen auch dort zu Spannungsänderung usw. Bei dieser gewöhnlichen Art der Erregungsweiterleitung wird die Information nicht durch die Stärke eines Aktionspotentials (der bewirkten Spannungsänderung) kodiert, sondern einzig durch die Häufigkeit der Aktionspotentiale in einer bestimmten Zeitspanne.
Die Erregungsweiterleitung in Stäbchen und Zapfen funktioniert jedoch auf andere Weise: Sie kodieren die Lichtinformation nicht über die Häufigkeit von Aktionspotentialen, sondern über das Ausmaß ihrer transmembranären Spannungsänderung. Die meisten anderen Neurone sind in ihrer Ruhelage (wenn kein Reiz eintrifft) mit etwa −65 mV negativ geladen. Wirkt auf sie ein Reiz ein, schnellt die Ladung für kurze Zeit auf etwa +10 bis +30 mV nach oben, und ein Aktionspotential wird durch diese Depolarisation ausgelöst. Stäbchen und Zapfen sind in ihrer Ruhelage (wenn kein Licht eintrifft) jedoch mit etwa −40 mV weniger stark negativ geladen – also leicht depolarisiert. Sobald Licht auf sie einwirkt, werden sie noch negativer geladen (bis max. etwa −65 mV) – also hyperpolarisiert – anstatt wie die anderen Neurone positiver zu werden. Grob gesagt, schüttet jedes Neuron umso mehr Botenstoffe aus, je positiver es geladen ist. Während normale Neurone also bei einem Reiz (der Depolarisation bewirkt) auf einmal sehr viel mehr Botenstoffe ausschütten, läuft diese Reaktion bei Fotorezeptoren genau umgekehrt ab: Trifft ein Lichtreiz ein, werden sie noch negativer (hyperpolarisiert) und schütten weniger Botenstoffe als in Ruhelage aus. Nachgeschalteten Zellen wird der Lichtreiz also nicht durch mehr, sondern durch weniger ausgeschüttete Botenstoffe signalisiert. Die Intensität des Lichtreizes wird den nachgeschalteten Zellen (Bipolarzellen) durch das Ausmaß der Botenstoffreduzierung mitgeteilt – je weniger Botenstoffe, desto stärker war der Lichtreiz.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 23.10. 2024