Transport (Biologie)
| Übergeordnet |
| Biologischer Prozess |
| Untergeordnet |
| Intrazellulärer Transport
Transmembrantransport Extrazellulärer Transport Sekretion Transepithelialer Transport |
| Gene Ontology |
|---|
|
|
Der Transport von Stoffen, Energie und Information ist für Lebewesen die Voraussetzung, ihren komplexen Stoffwechsel und andere Lebensvorgänge zu koordinieren und aufrechtzuerhalten.
Notwendigkeit eines Stofftransports
Alle Organismen stehen mit ihrer Umgebung in einem regen Stoffaustausch:
- Einerseits müssen sie Stoffe aus der Umgebung aufnehmen, die als Bau- oder Energiestoffe direkt oder nach Anpassung an die eigenen Bedürfnisse verwendet werden.
- Andererseits geben sie Stoffe an die Umgebung ab und verändern dadurch die Umwelt für sich und für andere Organismen (Exkretion von Gift- und Abfallstoffen, von Schleimstoffen, Schutzstoffen, Abwehrstoffen, Zersetzungsstoffen zum Auflösen von Mineralien oder organischem Material).
Aber auch innerhalb eines Organismus sind Stoffströme zu finden:
- Stoffe müssen vom Ort ihrer Entstehung (Syntheseort) zum Verbrauchs- oder Speicherort transportiert werden.
- Innerhalb einer Zelle findet auch ein Stoffaustausch zwischen den Zellorganellen und dem Cytosol statt.
Stoffe werden auch zwischen den Individuen einer Population (z.B. Pheromone) und zwischen den Populationen eines Ökosystems (z.B. Nahrung, Pollen) ausgetauscht.
In vielen Fällen sind die Stoffströme zu einem Kreislauf verknüpft (z.B. globaler Kohlenstoffkreislauf).
Zelluläre Transportsysteme
Diffusion: Die freie, ungehinderte Verteilung von Molekülen und Ionen in einem Raum hängt nur von der Temperatur und dem Konzentrationsgradienten ab. Sie tritt innerhalb einer Zelle oder außerhalb der Zelle in den sogenannten Interzellularen auf. Beispiele hierfür sind die Verteilung des Kohlenstoffdioxids in den Interzellularen des Schwammgewebes eines Blattes oder die Diffusion von Transmittermolekülen durch den synaptischen Spalt.
Zellwände, Zellmembranen und Organellmembranen stellen zunächst eine Diffusionsbarriere dar. Als Membrantransport wird der Austausch von Stoffen über diese Barriere hinweg bezeichnet. Permeation ist Diffusion durch diese Grenzschichten
Transmembran-Transport
Zellwände sind omnipermeabel, sie lassen alle Moleküle und Ionen, außer den Makromolekülen, passieren.
Biomembranen sind selektiv permeabel (Auslese der Stoffe), sie sind nur für kleine Moleküle wie Wasser, Kohlenstoffdioxid oder Sauerstoff durchlässig, die auf Grund von Unregelmäßigkeiten in der Lipid-Doppelschicht durch die Membran diffundieren. Da größere Moleküle wie Glucose oder Aminosäuren aber nur schwer die Membran passieren können, sind Transportmechanismen für eine erleichterte Aufnahme oder zum Aufbau eines Konzentrationsgradienten als Kurzzeit-Energiespeicher notwendig.
Im Unterschied zur Diffusion handelt es sich beim Proteintransport um einen enzymatischen Prozess, das heißt, da nur eine begrenzte Anzahl Transportproteine zur Verfügung steht, steigt die Prozessaktivität hyperbolisch an, bis sie einen Sättigungswert erreicht hat, der nicht überschritten werden kann.
Darüber hinaus sind diese Transportsysteme hoch selektiv, sie können bestimmte Moleküle aus der Umgebung erkennen und gezielt auswählen. Dabei können sie sogar Stereoisomere voneinander unterscheiden.
Den Transport durch eine Membran hindurch übernehmen in der Regel integrale Proteine, welche von einer Seite der Membran zur anderen reichen. Sie bilden einen Kanal, der innen hydrophil ist und auf Grund seines Durchmessers und bestimmter Ladung für bestimmte Moleküle oder Ionen selektiv ist.
Die Öffnung dieser Kanäle erfolgt meist erst auf Grund eines Signals (Hormone, Transmitter, Aktionspotential, mechanische Verformung), sodass der Transmembran-Transport nach Bedarf gesteuert werden kann.
Passiver Transport
Beim passiven Transport erfolgt die Wanderung von Molekülen oder Ionen auf Grund eines Konzentrationsgefälles, bei Ionen kann auch das Membranpotential noch eine Rolle spielen. Für den raschen Wassertransport ins innere der Zelle gibt es in der Zellmembran Aquaporine.
- Ionen-Kanäle:
- Sie erleichtern die Diffusion von Ionen durch die Membran entsprechend dem Konzentrationsgefälle und benötigen keine Energie. Beispiele: Natrium- und Kalium-Ionenkanäle der Nervenzellen, Calcium-Ionenkanäle der Nervenzellen und Muskeln, Chlorid-Kanäle von Nervenzellen und sekretorischen Zellen. Die Öffnung der Ionenkanäle wird entweder durch Änderung des Membranpotentials, durch Transmitter oder durch Ca2+ veranlasst.
- Carrier:
- Carrier (Trägerproteine) sind Transmembran-Proteine, die ähnlich wie Enzyme funktionieren: zunächst bindet sich das zu transportierende Molekül an eine spezifische Substratbindungsstelle an das Carrier-Protein, es bildet sich ein Carrier-Substrat-Komplex. Dieser Komplex ändert seine Konformation und das Molekül wird auf der anderen Seite der Membran wieder freigesetzt.
Cotransport: Manche Carrier tragen Bindungsstellen für verschiedene Substrate. Erst wenn sie alle besetzt sind, erfolgt der Transport. Symport ist der Transport aller Substrate in dieselbe Richtung, beim Antiport werden die Substrate in die entgegengesetzte Richtung transportiert. (Handelt es sich nur um ein Substrat, spricht man auch von Uniport.)
- Glucose-Transporter:
- Bei Bakterien und bei Säugern (Leber, β-Zellen der Bauchspeicheldrüse) wird der passive Transport durch das Kanal-Protein dadurch erleichtert, dass die Glucose im Inneren sofort zu Glucose-6-phosphat phosphoryliert wird, sodass sie nicht mehr durch den Kanal zurück diffundieren kann und die Glucose-Konzentration im inneren der Zelle niedrig bleibt. Das Glucose-6-phosphat kann in der Glykolyse sofort weiterverarbeitet werden. Das Phosphat stammt von Phosphoenolpyruvat (PEP) der Vorstufe von Brenztraubensäure, dem Endprodukt der Glykolyse.
Aktiver Transport
- aktiver Primärtransport – Ionenpumpen
- Sie erhöhen die Konzentrationsunterschiede zwischen den Kompartimenten einer Zelle, indem sie Ionen von der Seite der niedrigeren zur Seite der
höheren Konzentration transportieren. Die Energie entstammt der Hydrolyse von ATP zu ADP und Phosphat. Beispiele:
ATPasen (z.B.
Natrium-Kalium-Ionenpumpe)
Neben Ionen-Pumpen gibt es auch für Moleküle eigene ATP-abhängige Transport-Systeme. Beispiel: Transport von Peptiden in das endoplasmatische Retikulum.
- aktiver Sekundärtransport
- Wird die Energie des Konzentrationsgradienten nicht zum ATP-Aufbau, sondern zum Transport eines weiteren Ions genutzt, spricht man von Sekundärtransport. Auch hier gibt es Symport (die Stoffe werden in die gleiche Richtung transportiert) und Antiport (Transport in entgegengesetzte Richtung).
- Permeasen bei Bakterien
- Permeasen sind Kanalproteine, die aktiv Moleküle oder Ionen durch die Zellmembran transportieren. Sie sind ATP-unabhängig und entnehmen die Energie aus Konzentrationsgradienten oder dem Membranpotential.
| Art | Transportgut | Transportrichtung | treibende Kraft |
|---|---|---|---|
| Symport | Lactose und Protonen | nach innen | Membranpotential und pH-Gradient |
| Symport | Lactat und Protonen | nach innen | pH-Gradient |
| Antiport | Natrium-Kationen und Protonen | H+ nach innen, Na+ nach außen | pH-Gradient |
| Uniport | Lysin-Kationen | nach innen | Membranpotential |
Der Import von Glucose erfolgt durch das Transmembran-Enzym II C, welches Glucose beim Durchtritt phosphoryliert und damit die Konzentration der freien Glucose im inneren der Zelle niedrig hält. Der Maltose-Import durch die Maltose-Permease ist von ATP abhängig.
Vesikel-Systeme (Membranverlagernder Transport)
Makromoleküle und größere Nahrungspartikel können nicht mehr mittels Transportproteine durch die Membran hindurch treten. Sie werden mit Hilfe eines Vesikelsystems (lat. vesica, die Blase) transportiert. Im Inneren der Zelle werden diese Vesikel im Allgemeinen als Endosomen bezeichnet.
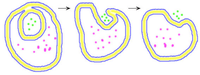
Vesikelbildung
|
Vesikel bilden sich dadurch, dass sich ein Teil der Membran ein- oder ausstülpt und sich dann als geschlossenes Hohlkügelchen abschnürt. Dabei werden Stoffe und Partikel des Milieus mit eingeschlossen. Umgekehrt kann ein Vesikel wieder mit der Membran verschmelzen und seinen Inhalt ausschütten.
| Export | Import |
|---|---|
 Exocytose |
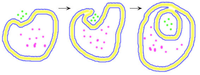 Endocytose |
 Abschnüren eines Vesikels |
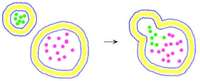 Verschmelzen eines Vesikels |
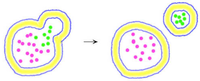
Bei der Endozytose (gr. kytos, die Zelle; gr. endon, innen) werden Stoffe oder Partikel in die Zelle importiert. Handelt es sich dabei um Nahrungspartikel, spricht man von Phagozytose (gr. phagos, Fresser). Wird Wasser mit darin gelösten Stoffen aufgenommen, wird das als Pinozytose (gr. pino, trinken, saugen) bezeichnet.
Bei der rezeptorvermittelten Endozytose werden nur bestimmte Moleküle zur Aufnahme ausgewählt. Beispiel: Cholesterin-Aufnahme. Bestimmte Stellen der Zellmembran sind außen mit Cholesterin-Rezeptoren und innen mit Eiweißen ausgekleidet (coated pits, Stachelsaum-Gruben). Sind alle Rezeptoren besetzt, stülpt sich die Membran ein, es bilden sich sogenannte Stachelsaum-Vesikel (coated vesicles).
Phagozyten sind spezialisierte Fresszellen des Immunsystems, die durch Phagozytose Zelltrümmer, Krankheitserreger und Fremdeiweiße, ja sogar Asbestfasern aufnehmen (siehe Makrophage).
Bei der Exozytose (gr. exo, außen, nach außen, außerhalb) werden Sekrete oder nicht verwertbare Stoffe ausgeschieden.
Beispiele:
- Amöben nehmen über Pseudopodien Nahrung auf.
- Ciliata strudeln durch Wimpernschlag Nahrungsteilchen in eine Empfangsvakuole. Diese löst sich vom Zellschlund ab. Während der Wanderung dieser Vakuole durch die Zelle, wird die Nahrung verdaut. Am Zellafter verschmilzt diese Vakuole wieder mit der Zellmembran und gibt dabei die unverdaulichen Bestandteile nach außen ab.
- Bei den Eukaryoten bilden Kernmembran und endoplasmatisches Retikulum (ER) einerseits und der Golgi-Apparat andererseits eigene Kompartimentsysteme, die untereinander über Vesikel miteinander in Verbindung stehen. So werden zum Beispiel im rauen ER Eiweiße synthetisiert, in Vesikel verpackt und zu den Diktyosomen des Golgi-Apparates transportiert. Dort verschmelzen die Vesikel mit den Golgi-Zisternen. In den Zisternen werden diese Eiweiße weiter verändert, wieder in Vesikel verpackt und an andere Zisternen weitergereicht. Als primäre Lysosomen enthalten die Golgi-Vesikel schließlich Verdauungsenzyme. Sie Verschmelzen mit den Vakuolen der Phagozytose, den Phagosomen, zu sekundären Lysosomen. Golgi-Vesikel mit Sekreten wandern zur Zellmembran, verschmelzen mit dieser und geben ihren Inhalt nach außen ab.
- Cytopempsis (gr. pempsis, Sendung) ist der Durchtritt von Phagosomen durch eine Zelle und ihre Weitergabe an die Nachbarzelle, ohne dass ihr Inhalt verändert wird. Sie tritt in den Epithelzellen der Gefäße und in den Epithelzellen des Darmes auf.
Cytoskelett
Das Cytoskelett besteht aus Mikrotubuli, Actin-Filamenten und intermediären Filamenten. Neben der Aufrechterhaltung und Änderung der Gestalt einer Zelle dient es vor allem innerhalb der Zelle dem Transport von Chromosomen bei der Zellteilung und dem Transport von Zellorganellen, Vesikeln und Makromolekülen.
Organismus
Zellverbindungen
- Zunächst verknüpfen diese Zell-Zell-Verbindungen Zellen zu einem Gewebe. In Abschlussgeweben dichten sie das Organ gegen die Außenwelt ab. Im Darmepithel ermöglichen sie aber auch die Resorption von Wasser und Ionen.
- Gap-Junctions treten nur bei den Eumetazoa auf.
Um den Austausch von Stoffen zwischen den Zellen eines Gewebes zu ermöglichen, wird die Lücke (engl. gap) zwischen den Zellen durch Poren von 1,2 bis 2 nm Durchmesser, überbrückt (engl. junction Verbindung). Diese Poren werden von 6 radial angeordneten Proteinen gebildet, sie befinden sich in den Membranen beider Zellen und stehen sich genau gegenüber, sodass ein Kanal durch beide Zellmembranen entsteht. Sie sind omnipermeabel und lassen sowohl Ionen als auch ungeladene Moleküle entsprechend dem Konzentrationsgefälle passieren. Sie können aber auch selektiv permeabel sein und im Herz- und Nervengewebe als elektrische Synapsen die Weiterleitung von Aktionspotential ermöglichen.
- In den Zellwänden benachbarter Pflanzenzellen sind Poren mit einem Durchmesser von 20 bis 40 nm zu finden, über die ein direkter Kontakt zwischen dem Cytosol der beiden Zellen besteht. Auf die Weise bildet das Cytosol aller Zellen eines Pflanzengewebes eine zusammenhängende Einheit, den Symplast. Die Plasmodesmen werden von Desmotubuli, massiven Proteinen, die das endoplasmatische Retikulum der beiden Zellen zu einer Einheit verbinden, durchzogen. Besonders viele Plasmodesmen enthalten die Querwände der Siebröhre, Leitungsbahnen der Pflanzen für Assimilate, und die Tüpfel der Pflanzenzellen.
- Bei Pflanzen erfolgt der Nahtransport von Wasser und darin gelösten Nährsalzen und organischen Stoffen in den Wurzeln über den Apoplast, das zusammenhängende System der Interzellularen und Hohlräume der Zellwände, durch Diffusion. Da Wurzeln aktiv Ionen aus der Umgebung aufnehmen, erhöht sich die Konzentration im Apoplasten und Wasser strömt durch Osmose nach. In der Endodermis sind die Casparyschen Streifen durch Suberin-Einlagerungen in den Zellwänden wasserundurchlässig. Hier werden Ionen aktiv und selektiv in den Symplasten transportiert.
Geißeln und Wimpern
Mit Hilfe von Geißeln und Wimpern (Zilien) können Strömungen erzeugt werden, mit deren Hilfe Nahrungspartikelchen an den Ort der Phagocytose transportiert werden.
Beispiele:
- Bei der Kragengeißelzelle Monosiga ist die Basis der Geißel von einem Kragen feiner Stäbchen, die von der Zellmembran gebildet werden, umgeben. Dieser Stäbchenkranz funktioniert wie eine Reuse: Der Geißelschlag erzeugt eine Wasserströmung durch den Kragen hindurch. Nahrungspartikel bleiben an den mit Schleim überzogenen Kragenstäbchen hängen und wandern mit dem Schleim zur Basis des Kragens, wo sie phagozytiert werden. Kragengeißelzellen finden sich als spezialisierte Zellen auch bei den Schwämmen. Hier entsteht auch ein beständiger Wasserstrom von außen durch Poren in den Zentralraum und von dort durch die obere Öffnung wieder nach außen.
- Bei dem Pantoffeltierchen Paramecium sorgen Wimpern im Mundfeld für den Transport von Nahrungspartikeln in die Empfangsvakuole.
- Die Schleimhaut der Atemwege wird von zwei Zelltypen gebildet: Becherzellen produzieren einen Schleimfilm, in dem Staubpartikel und Krankheitserreger hängen bleiben. Dieser Film wird von den Zellen mit Zilien Richtung Mund- und Nasenhöhle transportiert wird.
- Der Transport der Eizelle im Eileiter erfolgt durch dessen Flimmerepithel.
Ferntransport – Organsysteme
Je größer ein vielzelliger Organismus ist, umso schlechter werden im Inneren liegende Zellen allein durch Diffusion und zelluläre Transportmechanismen versorgt. Eigene Transporteinrichtungen bewerkstelligen eine rasche und gleichmäßige Verteilung von Stoffen und Wärme. Je größer der Organismus ist, umso stärker verzweigt sind diese Transportsysteme.
Der Transport muss unabhängig von der Schwerkraft in jeder Körperlage möglich sein.
Tiere
Verdauungstrakt
Mit zunehmender Größe der Nahrungsteilchen ist es notwendig, diese mechanisch zu zerkleinern und chemisch bei den größeren Mehrzellern der Verdauungstrakt. Der Weitertransport der Nahrung erfolgt über Längs- und Ringmuskeln durch Peristaltik. Die durch Verdauung erschlossenen Nährstoffbausteine und andere kleinere Moleküle diffundieren durch die Darmwand oder werden durch eigene Transportproteine durch die Zellmembranen in die Körperhöhle geschleust.
Blutkreislauf
Blut und Lymphe transportieren zahlreicher Stoffe (Nährstoffe, Abwehrstoffe, Abfallstoffe, Hormone) und Wärme. Als Transportmittel dient Wasser, in dem die Stoffe entweder gelöst oder an Trägermoleküle gebunden sind.
Das Blut wird durch ein spezielles Organ, das Herz, durch den Körper gepumpt. Bei den Wirbeltieren unterstützen Gefäßmuskeln und angrenzende Skelettmuskeln diese Pumptätigkeit.
Insekten und Weichtiere besitzen einen offenen Blutkreislauf, Wirbeltiere und Ringelwürmer einen geschlossenen. Dabei wird das Blut in einem geschlossenen Gefäßsystem durch den Körper geführt. Bei den Organen verzweigen sich die Blutgefäße zu engen, dünnwandigen Kapillaren. Hier findet der Stoffaustausch mit dem angrenzenden Gewebe durch Diffusion statt.
Gastransport
Der Gasaustausch zwischen Umwelt und Organismus findet entweder über die Haut (Amphibien und im Wasser lebende Würmer), über Kieme (Fische, Amphibien-Larven, Wasserschnecken, Krebse) oder über Lunge (an Land lebende Tiere) statt.
Um die Kiemen immer mit frischem Wasser zu versorgen, haben sich verschiedene Mechanismen entwickelt: Fische saugen über das Maul frisches Wasser an und stoßen es über die Kiemen wieder aus, Krebse erzeugen mit ihren Hinterleibsbeinen einen beständigen Wasserstrom, die sessilen Röhrenwürmer bewegen ihre Kiemen durchs Wasser.
Um Lungen mit Frischluft zu versorgen, wird durch Erweiterung des Brustraumes mittels Zwerchfell und Zwischenrippenmuskulatur ein Unterdruck erzeugt, der die Luft einsaugt. Durch Verengung des Brustraumes wird die mit Kohlenstoffdioxid angereicherte Luft wieder ausgepresst.
Transport der Atemgase im Körper:
- Sauerstoff (Mensch): Der Sauerstoff diffundiert durch die Lungenbläschen ins Blut. Nur 3 % lösen sich physikalisch, die übrigen 97 % werden an das Hämoglobin der Erythrozyten gebunden und so im ganzen Körper verteilt. In den Kapillaren wird der Sauerstoff wieder abgegeben, in den Muskeln wird er vom Myoglobin übernommen.
- Kohlenstoffdioxid (Mensch): Durch Diffusion gelangt das Kohlenstoffdioxid ins Blut, wird dort teilweise physikalisch gelöst (10 %), reagiert mit Wasser zu Hydrogencarbonat und verbleibt im Plasma (45 %) oder wird von den Erythrozyten als Hydrogencarbonat (35 %) oder an Hämoglobin gebunden (10 %) transportiert.
- Tracheensystem: Da das offene Blutkreislaufsystem der Insekten die Atemgase für den hohen Bedarf der Flugmuskulatur nicht schnell genug transportieren würde, haben sie ein eigenes Röhrensystem für den Gastransport, das Tracheensystem. Die Ventilation erfolgt durch Kontraktion und Erweiterung des Hinterleibs.
Weitere Transportsysteme
Weitere Transportsysteme sind die Nierenkanälchen, Harnleiter, Samenleiter, Gallengänge, Milchgänge der Milchdrüsen und Ausführgänge der Bauchspeicheldrüse, der Talgdrüsen und der Schweißdrüsen der Haut.
Nervensystem
Das Nervensystem der Tiere stellt ein Organsystem zur Aufnahme, Weiterleitung, Verarbeitung und Speicherung von Information dar. Grundlage ist der Transport von Ionen durch die Membran der Nervenzellen und von Transmitter-Stoffen durch den synaptischen Spalt. Auch Pflanzenzellen sind befähigt, Aktionspotentiale auszubilden. Ihre Entstehung dauert allerdings länger und wird nicht über ein eigenes Nervensystem weitergeleitet.
Pflanzen
Der Ferntransport von Wasser und darin gelösten Nährsalzen findet in den Tracheen und Tracheiden des Xylems statt, Assimilate werden in den Siebröhren des Phloems transportiert.
- Transpirationsstrom
- Im Transpirationsstrom werden Wasser und darin gelöste Ionen sowie organische Verbindungen und Alkaloide des Wurzelstoffwechsels durch die Gefäße des Xylems nach oben transportiert.
- Da die Gefäße sehr eng sind, sorgt die Kapillarkraft dafür, dass das Wasser bis zu 80 Zentimeter aufsteigen kann. Der Wurzeldruck ist eine Folge des aktiven, ATP verbrauchenden, Ionentransports in den Wurzeln, er erlaubt einen Anstieg der Wassersäule bis auf 10 Meter. Der Transpirationssog entsteht durch das Verdunsten von Wasser über die Blätter. Da eine durch Kohäsion zusammenhängende, bis zu den Wurzeln reichende, Wassersäule besteht, die durch Adhäsion an den Gefäßwänden haftet, kann das Wasser bis auf eine Höhe von 120 Meter gehoben werden.
- Die Assimilate (Mono- und Disaccharide) werden in den Siebröhren nach unten transportiert. Ihr Zytoplasma bildet einen zusammenhängenden Symplasten, da ihre Querwände (die Siebplatten) zahlreiche Poren aufweisen, die von Plasmodesmen durchzogen sind. Der Transport erfolgt entlang eines starken osmotischen Gefälles. Die Siebröhrenzellen erhalten die Assimilate durch aktiven, ATP-verbrauchenden Transmembran-Transport von den sie umgebenden Geleitzellen (Transferzellen). Ihr Plasmalemma ist zur Oberflächenvergrößerung stark gefaltet (ähnlich den Darmzotten), sodass viele Transportproteine Platz haben. Am Verbrauchs- oder Speicherort werden die Assimilate wieder aktiv aus den Siebröhren gepumpt.
- Gastransport
- Der Gastransport erfolgt bei Pflanzen allein durch Diffusion über Spaltöffnungen oder Lentizellen und das die ganze Pflanze durchziehende Interzellular-System. Besonders ausgeprägt sind diese Hohlräume im Schwammgewebe des Blattes. Bei Sumpf- und Wasserpflanzen wird der Gasaustausch besonders in den untergetauchten Pflanzenteile durch Aerenchyme (Durchlüftungsgewebe, Sternparenchym bei Binsen) und Lacunen gefördert. Bei Sumpfpflanzen (z.B. Mangroven) verbessern Atemwurzeln den Gasaustausch mit der Luft.
Ökosysteme
Energiefluss
Der Aufbau und die Aufrechterhaltung von Strukturen macht bei den Lebewesen eine beständige Aufnahme von Energie notwendig. Über die Nahrungsketten wird diese Energie in Form von energiereichen Nährstoffen weitergegeben. Diese Energie stammt letztlich aus der Sonnenstrahlung, die von phototrophen Organismen genutzt und als chemische Energie in Nährstoffen gespeichert wird. Bei jedem Stoffwechselprozess geht Energie in Form von Wärmebewegung verloren.
Stofffluss
Innerhalb eines Ökosystems findet ein ständiger Austausch von Nährstoffen, Mineralstoffen und Gasen (O2, CO2) statt, die in einem Kreislauf geführt werden.
Die Primärproduzenten (photoautrotrophe Pflanzen, eukaryotische Einzeller und chemoautotrophe Bakterien) bauen bei der Assimilation (Biologie) aus anorganischen Stoffen organische Stoffe auf, die sie selbst als Bau- und Energiestoffe nutzen. Die organischen Stoffe (echte Nährstoffe: Proteine, Kohlenhydrate, Fette, DNA) sind somit Träger von Energie und Struktur des Lebens, die bei der Assimilation aus den energiearmen anorganischen Stoffen (Mineralsalze, CO2, H2O, fälschlicherweise auch bezeichnet als „Nährstoffe“ der Pflanzen) aufgebaut werden. Destruenten bauen die organische Substanz wieder ab und führen sie so in den anorganischen Zustand zurück: der Kreislauf ist geschlossen. Tiere nehmen als heterotrophe Konsumenten an dem Stoffkreislauf teil, indem sie organische Nährstoffe aufnehmen, zum Teil veratmen und in anorganische Stoffe zurückverwandeln oder an die Destruenten oder weitere Konsumenten weitergeben.
Ein autarkes Ökosystem könnte also auch ohne Tiere als Konsumenten existieren, nicht aber ohne Destruenten. Die Rolle der Tiere beim Stofffluss im Ökosystem beruht auf der raschen Zerkleinerung organischen Materials bei der Ernährung, sodass Pilze und Bakterien als Destruenten die Remineralisierung rascher durchführen können.
Beispiele:
 |
 |
 |
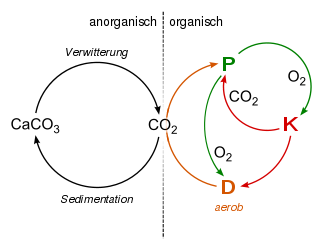
| Kohlenstoff-Sauerstoff-Kreislauf unter aeroben Bedingungen:
Dieser biotische Kreislauf ist mit dem abiotischen Kohlenstoffzyklus gekoppelt: Ein Teil des CO2 sedimentiert im Wasser als Calciumcarbonat. Kalkgestein verwittert wieder. |
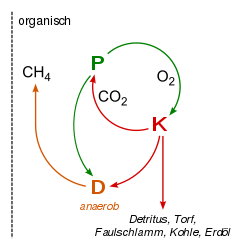
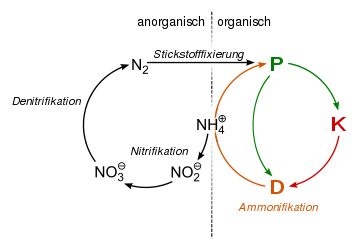
Kohlenstoff-Sauerstoff-Kreislauf unter anaeroben Bedingungen. | Stickstoffkreislauf: Stickstofffixierung, Ammonifikation, Nitrifikation und Denitrifikation erfolgt durch Bakterien. Die Nitrifikanten sind streng aerob, die Denitrifikation erfolgt unter anaeroben Bedingungen. |
Legende:
| ||
Symbiosen
In Symbiosen werde Stoffe zwischen den Partnern besonders effektiv ausgetauscht. Man unterscheidet Ektosymbiosen und Endosymbiosen. Bei diesen lebt der eine Partner im Inneren des anderen. Beispiele hierfür sind die Stoffwechselsymbiosen der Korallen und Radiolarien mit Grünalgen oder Cyanobakterien: Der photosynthetisch aktive Partner erhält vom Tier Kohlenstoffdioxid aus der Zellatmung. Durch die Photosynthese entsteht Sauerstoff, der dem Tier für die Dissimilation zur Verfügung steht. Zusätzlich assimilieren die autotrophen Partner Stickstoff und geben die stickstoffhaltigen, organischen Verbindungen an das Tier weiter. Grünalgen nutzen Nitrat als Stickstoffquelle, Cyanobakterien den elementaren Stickstoff.
Der Stoffkreislauf kann auch gestört sein
- Unter anaeroben Bedingungen ist der Abbau langsamer. So kann sich totes, organisches Material ansammeln, das sich in geologischen Zeiträumen zu Torf, Kohle oder Erdöl umwandelt – die darin gebundenen anorganischen Ausgangsstoffe (v.a. CO2) sind so dem Kreislauf entzogen. Demzufolge hat sich das bei der Produktion gebildete O2-Gas in der Atmosphäre stark angereichert. Unser heutiger O2-Gehalt der Atmosphäre stammt also aus der Fossilisation toter Biomasse, und bei der Verbrennung dieser Stoffe wird wieder nicht nur CO2 gebildet, sondern auch O2 verbraucht.
- Durch Wasserströme werden ständig gelöste Mineralstoffe zwischen Ökosystemen ausgetauscht. Vor allem die intensive Düngung in der Landwirtschaft trägt zu diesen Strömen bei und sorgt für Störungen von ökologischen Stoffkreisläufen.
Geophysikalischer Transport
Materialien und Stoffe, die den Lebewesen als Lebensgrundlage dienen können, werden durch geophysikalische Vorgänge einem Ökosystem zugeführt oder entnommen.
Zirkulation im See
Ein stehendes Gewässer erwärmt sich durch die Sonneneinstrahlung von oben. Ist es tief genug wie bei einem See, entsteht eine ausgeprägte Temperaturschichtung. Das warme Oberflächenwasser (Epilimnion) schwimmt auf dem kalten Tiefenwasser (Hypolimnion). Die beiden Wasserkörper sind durch die Sprungschicht (Metalimnion mit Thermokline) voneinander getrennt. Ist die Schichtung stabil, kann durch den Wind nur das Epilimnion durchmischt und mit Sauerstoff für die Tiere oder Kohlenstoffdioxid für photoautotrophe Organismen aus der Atmosphäre versorgt werden. Totes organisches Material sinkt zu Boden und wird dort durch aerobe Destruenten remineralisiert. Da aber kein Austausch zwischen Hypo- und Epilimnion stattfindet, reichern sich die Nährsalze unten an, während die obere Schicht an Nährsalzen verarmt, wodurch dort der Zuwachs an Primärproduzenten eingeschränkt wird.
Kühlt das Oberflächenwasser auf die Temperatur des Tiefenwassers ab, bricht die Sprungschicht zusammen, der gesamte Wasserkörper kann durchmischt werden. Die Nährsalze werden durch Konvektion in die oberen Wasserschichten transportiert und stehen damit wieder den photoautotrophen Organismen zur Verfügung, das an Sauerstoff reiche Oberflächenwasser wird nach unten transportiert, somit steht wieder Sauerstoff den Destruenten und Tieren des Tiefenwassers zur Verfügung.
Zirkulation im Meer
Ähnlich wie bei einem See bildet sich auch in den Meeren eine Sprungschicht aus, die nur eine Durchmischung des Oberflächenwassers erlaubt. Beständige Winde, Corioliskraft, Gezeiten sowie Temperatur- und Salzgehaltsgefälle erzeugen ein lokales und globales, horizontales und vertikales, über die Sprungschicht hinausgreifendes, Strömungssystem. Der Golfstrom ist Bestandteil dieses Systems. Er transportiert an der Oberfläche warmes Wasser nach Norden und sorgt damit in diesen Breiten für ein gemäßigtes Klima. Die in der Sargassosee geschlüpften Aal-Larven werden durch den Golfstrom nach Nord- und Westeuropa transportiert, wo sie flussaufwärts zu ihren Laichgebieten schwimmen.
Durch die Verdunstung erhöht sich der Salzgehalt des Golfstroms und im Norden wird das Wasser abgekühlt. Dadurch erhöht sich die Dichte des Wassers und es sinkt östlich von Grönland nach unten ab. Als kaltes, salzreiches Tiefenwasser strömt es wieder nach Süden und reichert sich dabei mit Nährsalzen an. In den Auftriebszonen des Peru-, Benguela- und Kanarenstroms sorgen diese Nährsalze für eine hohe Biomasseproduktion.
Das verstärkte Abschmelzen der Eiskappen des Nordpols und Grönlands würden bei einer Klimaerwärmung zu einer Erniedrigung des Salzgehaltes und damit zum Abreißen des Golfstromes führen, dies würde zu einer eiszeitartigen Abkühlung in Europa führen.
Verfrachtungen durch Flüsse
Im Gegensatz zu einem See stellt ein Fluss ein offenes Ökosystem dar: Durch die Erosion im Oberlauf reichert sich das Wasser mit Nährsalzen (Carbonate, Sulfate, Eisenionen) an, die flussabwärts transportiert werden und von den Primärproduzenten und Konsumenten genutzt werden. Im Mittel- und Unterlauf kommt es einerseits am Prallhang zu Abtragungen im Uferbereich, dadurch gelangt organisches Material (Detritus) in den Fluss, das von Destruenten remineralisiert wird und damit den Produzenten zur Verfügung steht. Andererseits kommt es zu Aufschüttungen von Sand- und Schlickbänken, die besondere Kleinbiotope darstellen. Deltagebiete können durch Aufschüttungen einen Zugewinn an Land bewirken (Euphrat und Tigris, Mississippi River). Totes organisches Material von den Lebewesen des Flusses oder des Uferbereichs gelangt in großen Mengen in den Unterlauf, sodass dieser Bereich ein nährstoffreiches (eutrophes) Ökosystem darstellt.
Regelmäßige (Nil) oder gelegentliche Überschwemmungen (Oderbruch) versorgen das Überschwemmungsgebiet mit anorganischen und organischen Nährstoffen und schaffen damit die Grundlage für eine erhöhte Biomasseproduktion an Land.
Windverfrachtungen
Ähnlich wie bei den Flüssen kommt es auch durch den Wind zur großräumigen Verfrachtung von organischem und anorganischem Material. Dadurch kann Humus in einem Gebiet verloren gehen und in anderen Gebieten wieder angereichert werden.
Lebewesen nutzen den Wind als Transportmittel: Windbestäuber (Fichte, Tanne, Kiefer, Gräser) lassen ihren Pollen transportieren. Pilze nutzen den Wind zur Verbreitung ihrer Sporen. Viele Blütenpflanzen lassen ihren Samen als Flugfrüchte durch den Wind verbreiten (Löwenzahn, Weide, Ahorn). Jungspinnen lassen sich auf einem Faden sitzend durch den Wind in andere Regionen transportieren.
Die Verfrachtung von flugfähigen Tieren kann ebenfalls zur Verbreitung ihrer Art führen (Besiedlung der pazifischen Inselwelt durch Vögel und Insekten, Wanderrichtung der Wanderheuschrecke). Allerdings können diese Tiere auch aufs Meer hinaus geweht werden, was zu ihrem Tod führen kann. Deshalb findet man auf kleinen Inseln häufig flugunfähige Insekten oder Vögel (Dronte).
Tiere als Transportsysteme
Auch Tiere werden von anderen Lebewesen als Transportsysteme genutzt:
- Tierbestäubung: Insekten (Biene, Hummel), Vögel (Kolibri), Fledermäuse, Primaten transportieren den Pollen von einer Blüte zur anderen.
- Elterntiere transportieren Jungtiere im Maul an einen sicheren Ort.
- Krötenmännchen lassen sich längere Zeit während der Laichzeit von den Weibchen tragen.
Literatur
- Anita Roth-Nebelsick: Die Prinzipien der pflanzlichen Wasserleitung. In: Biologie in unserer Zeit. 36(2), S. 110–118 (2006), ISSN 0045-205X.
Siehe auch


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 20.06. 2026