Vielzellige Tiere
| Vielzellige Tiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Oben: Schwämme,
Seeanemone,
Regenwurm | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Metazoa | ||||||||||||
| Haeckel, 1874 |
Die vielzelligen Tiere (wissenschaftlich Metazoa, von altgriechisch μετα meta, deutsch ‚danach‘ und ζῷον zóon ‚Tier‘) sind ein zoologisches Taxon, in dem alle mehrzelligen Tiergruppen zusammengefasst werden. Heute sind über 1,3 Millionen Arten von vielzelligen Tieren bekannt. Mit den noch nicht beschriebenen wird geschätzt, dass es 10 bis 20 Millionen Arten sind. Von den bisher bekannten Arten machen die Gliederfüßer (Arthropoda) mit etwa einer Million über 80 % aus, und innerhalb der Gliederfüßer stellen die Käfer und die Schmetterlinge zusammen die Hälfte der Arten. Gliederfüßer und Weichtiere, mit 100.000 Arten der zweitartenreichste Stamm, umfassen 90 % der heute lebenden Arten. Die Wirbeltiere stellen 5 % des Artenreichtums der Tierwelt.
In wissenschaftlichen Veröffentlichungen werden die Metazoa heute oft synonym zu den Tieren (Animalia) verwendet, für die Metazoa und die einzelligen Vertreter aus ihrer Stammgruppe wurde 2002 das neue Taxon Holozoa gebildet.
Merkmale
Kennzeichen der Metazoa ist neben dem Umstand, dass sie aus mehreren Zellen aufgebaut sind, auch die Differenzierung und Spezialisierung dieser Zellen. So finden sich bereits bei den Schwämmen unterschiedliche Zelltypen, die beispielsweise dem Nahrungserwerb, der Atmung (Kragengeißelzellen) oder dem Skelettaufbau dienen. Weitere Kennzeichen sind eine spezialisierte Gametengenese, begeißelte Spermien, die Furchung (Zellteilungen bei der befruchteten Eizelle durch Abschnürung) sowie das Genom der Mitochondrien.
Differenziertes Gewebe
Bedeutendstes Kennzeichen der Metazoa ist der vielzellige Körper, der aber keine Kolonie gleicher Zellen ist, die mehr oder weniger auch unabhängig voneinander existieren könnten. Die Zellen sind spezialisiert und morphologisch und funktionell differenziert. Gleichartige Zellen kommunizieren mittels chemischer Signale untereinander, beeinflussen sich gegenseitig, arbeiten zusammen und bilden Gewebe. Die Kommunikation dient unter anderem dem Erhalt der Körperform und der Form der Organe.
Gewebegrundtypen sind das Epithel (Auskleidungsgewebe) und das Bindegewebe. Das Epithelgewebe besteht aus einem geschlossenen Zellverband, fast ohne interzelluläre Zwischenräume. Die Zellen sind in Funktion und Struktur oft asymmetrisch (polarisiert) und auf unterschiedliche Weise miteinander verbunden.
Bindegewebe sind dagegen lockerer aufgebaut, mit isolierten Zellen in der extrazellulären Matrix. Die extrazelluläre Matrix besteht aus netzartig angeordneten Kollagenfasern, Wasser und Glykoproteinen. Epithel und Bindegewebe sind durch die Basalmembran voneinander getrennt.
Cilien
Die Cilien der Spermien und anderer Metazoenzellen haben quergestreifte Wurzeln und senkrechte Zentriolen, von denen eines den Basalkörper der Cilie bildet. Ansonsten weisen sie den einheitlichen Aufbau aller Eukaryoten-Cilien auf.
Gametogenese
Zur geschlechtlichen Fortpflanzung produzieren mehrzellige Tiere Eizellen und Spermien. Ihre Gametogenese ist ähnlich. Aus einer diploiden Mutterzelle entstehen vier haploide Zellen, entweder vier Spermien oder die befruchtungsfähige Eizelle und drei Polzellen, die normalerweise schnell wieder vergehen. Es wird angenommen, dass die Produktion einer großen, mit Nährstoffen versehenen Eizelle zu aufwendig ist, als dass aus jeder der vier Zellen eine Eizelle entstehen kann.
Furchung
Nach der Befruchtung teilt sich die entstandene diploide Zygote zunächst, ohne dass ein Zellwachstum stattfindet, in viele kleine Tochterzellen (Blastomere). Bei dieser Furchung genannten Zellteilung entstehen die Grundachsen des künftigen Embryos und eine erste Differenzierung. Aus der Furchung entsteht die Blastula, die sich in der Gastrulation einstülpt. Im Zuge der weiteren Differenzierung teilen sich die Zellen teilweise asymmetrisch, aus einer Stammzelle entsteht eine Stammzelle und eine Zelle, die sich weiter differenziert.
Die Arten der Furchung dienten lange als Merkmal, um die verschiedenen Tierstämme verwandtschaftlich einander zuzuordnen. Allerdings haben viele Stämme einen nur ihnen eigenen Furchungstyp entwickelt, und die systematische Bedeutung der verschiedenen Furchungstypen ist nur schwer zu beurteilen.
Mitochondrien
Die Mitochondrien der Metazoa haben ein vergleichsweise kleines Genom mit etwa 16.000 Nukleotiden auf 37 Genen. Im Vergleich dazu ist das Genom der Choanoflagellaten viermal so lang. Zudem weisen die Transfer-RNA und die ribosomale RNA eine ungewöhnliche Struktur auf.
Systematik
Das Schwestertaxon der vielzelligen Tiere – also der eigentlichen Tiere im modernen Wortsinn (Metazoa) – besteht aus den Kragengeißeltierchen (Choanoflagellata). Tiere und Kragengeißeltierchen bilden gemeinsam die Gruppe der Choanozoa. Sie werden mit den Einzellern der Filasterea zusammengefasst zur Abstammungsgemeinschaft der Filozoa, die gemeinsamen mit den Einzellern der Teretosporea die Gruppe der Holozoa formen. Das Schwestertaxon der Holozoa besteht aus den Nucletmycea, denen zum Beispiel die Pilze (Fungi) zugeordnet werden. Sie bilden die zwei Zweige der Schubgeißler (Opisthokonta).
Äußere Systematik der Metazoa
| |
| |
Die basalsten vielzelligen Tiere bestehen aus dem Monophylum der Schwämme (Porifera). Ihr Schwestertaxon wird Epitheliozoa genannt, in dem alle übrigen Tiere geordnet werden. Das Taxon enthält die Scheibentiere (Placozoa) und die Gewebetiere (Eumetazoa). Letztere teilen sich in die Gruppen der Hohltiere (Coelenterata) und der Zweiseitentiere (Bilateria). Die Hohltiere umfassen Rippenquallen (Ctenophora) und Nesseltiere (Cnidaria). Die Zweiseitentiere gliedern sich in Xenacoelomorpha und Nierentiere (Nephrozoa). Letztere bestehen aus Urmundtieren (Protostomia) und Neumundtieren (Deuterostomia).
| Innere Systematik der Metazoa | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Die eben dargestellte innere Systematik der Tiere birgt in sich das Coelenterata-Konzept. Eine Klade von Hohltieren aus Rippenquallen und Nesseltieren wird als Schwestertaxon der Zweiseitentiere gezeigt. Mit der Abbildung könnte eine taxonomische Sicherheit suggeriert werden, die allerdings zum gegenwärtigen Stand der Forschung nicht existiert. Tatsächlich werden die genauen verwandtschaftlichen Beziehungen zwischen den monophyletischen Gruppen der Schwämme, Scheibentiere, Rippenquallen, Nesseltiere und Zweiseitentiere diskutiert. Das Acrosomata-Konzept postuliert eine Gruppe aus Rippenquallen und Zweiseitentieren namens Acrosomata. Das Planulozoa-Konzept sieht eine Abstammungsgemeinschaft in Nesseltieren und Zweiseitentieren mit der Bezeichnung Planulozoa. Das Planulozoa-Konzept geht auf in dem Parahoxozoa-Konzept, bei dem die Scheibentiere zusammen mit den Planulozoa eine Klade mit dem Namen Parahoxozoa formen. Diese Tiere werden wiederum mit den Schwämmen zu den Benthozoa zusammengefasst, die das Schwestertaxon zu den Rippenquallen bilden.
Konzepte zur Systematik der Metazoa
| ||
| Coelenterata-Konzept | Acrosomata-Konzept | ParaHoxozoa/Planulozoa-Konzept |
|---|---|---|
|
|
|
Stammesgeschichte

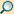
Der zeitliche Ursprung der Metazoa ist bis heute unsicher und umstritten. Dies liegt daran, dass alle Fossilien aus dem Präkambrium, insbesondere aber alle, für die ein Alter vor dem Ediacarium angegeben worden ist, schwer interpretierbar und in ihrer Zuordnung umstritten sind. Zahlreiche Angaben älterer Fossilien wurden von anderen Forschern später in Zweifel gezogen oder sogar nachträglich zweifelsfrei als Reste von Metazoa widerlegt. Altersabschätzungen anhand der Methodik der molekularen Uhr sind eher noch unzuverlässiger, da sie zur Kalibrierung auf sicher datierten Funden oder Ereignissen beruhen müssen, die nicht abgesichert vorliegen. Zahlreiche Datierungen mit dieser Methode ergaben daher bei weitem zu hohe Altersschätzungen. Auf bestimmten Molekülen in sehr alten Sedimenten, die als Biomarker für bestimmte Gruppen, vor allem für Schwämme, herangezogen worden sind, beruhende Datierungen sind sehr umstritten, da diese Verbindungen anders entstanden sein könnten (oder möglicherweise nur als Verunreinigung in die zu datierende Schichten eingedrungen sind). Interpretationen anhand des Verzweigungsmusters der basalen Kladen der Vielzelligen Tiere, die letzte der möglichen Methoden der Datierung, sind problematisch, weil es keinen wissenschaftlichen Konsens darüber gibt, welche der Gruppen tatsächlich die älteste (d.h. diejenige, die zuerst von der gemeinsamen Stammgruppe abzweigte) ist.
Unklar ist schon, welche Merkmale ein möglicher fossiler Stammgruppen-Vertreter der Metazoa aufgewiesen haben könnte. Peter Ax gibt als Autapomorphien der Metazoa folgende Merkmale an: Getrenntgeschlechtlicher („gonochorischer“) Organismus aus diploiden somatischen Zellen, haploiden Gameten und diploider Zygote, Oogenese mit einer Eizelle und drei rudimentären Polkörpern, Spermium differenziert in einen Kopf mit dem Zellkern und einer Geißel, mit einem Mittelstück („Hals“ mit Zentrosom mit zwei rechtwinklig angeordneten Zentriolen und vier Mitochondrien), bei der Spermatogenese entstanden als vier identische Spermien aus jedem Spermatozyten. Das Gewebe der somatischen Zellen ist durch dichte Zell-Zell-Verbindungen (Tight Junctions) für bestimmte Stoffe nur selektiv durchlässig, was ein inneres Milieu ermöglicht. Es ist eine extrazelluläre Matrix vorhanden, die Fasern aus Kollagen enthält. Die Entwicklung aus der Keimzelle erfolgt durch radiale Furchung. Für das ausgewachsene Tier nimmt er eine abgeflachte Hohlkugel (Blastula, Placula) als Organisationsform an, die aus begeißelten Zellen auf der Außenseite zur Fortbewegung und Nahrungsaufnahme und unbegeißelten Zellen auf der Innenseite besteht.
Von den genannten Merkmalen ist, mit Einschränkungen, möglicherweise ein einziges, die Radiärfurchung, an Fossilien tatsächlich zu beobachten. Fossilien von Gebilden, die als Zellen mit radiärer Furchung (mögliche Embryonen von Vielzellern) interpretiert werden, liegen aus der Doushantuo-Formation aus Südchina vor, die sogenannten Weng'an Biota, deren Alter auf etwa 570 bis 600 Millionen Jahre datiert wird. Aus derselben Formation liegen auch Fossilien anderer möglicher vielzelliger Tiere vor, deren Interpretation aber umstritten ist. Dabei handelt es sich vermutlich um die ältesten überzeugenden fossilen Nachweise von Metazoa. Noch ältere Fossilien (oder auch Spurenfossilien) wurden zwar in größerer Zahl vorgeschlagen, doch konnten die meisten davon als Fossilien vielzelliger Algen oder Zellfäden von Prokaryoten (wie etwa Cyanobakterien) identifiziert werden, die übrigen sind äußerst zweifelhaft und umstritten.
In den jüngeren Schichten des Ediacariums, nach der Gaskiers-Eiszeit vor etwa 580 Millionen Jahren, finden sich die Fossilien der rätselhaften Ediacara-Fauna. Diese sind in der Interpretation rundweg umstritten, doch werden einige der berühmten Fossilien wie Kimberella, Dickinsonia oder Eoandromeda von den meisten Forschern als Metazoa akzeptiert. Die ältesten Vertreter der heute lebenden Tierstämme, deren Zuordnung unzweifelhaft gesichert ist, stammen aber tatsächlich erst aus dem Kambrium vor etwa 541 Millionen Jahren. In der unterkambrischen chinesischen Chengjiang-Faunengemeinschaft, vor etwa 520 Millionen Jahren, sind bereits nahezu alle Tierstämme, die überhaupt das Potenzial zur Fossilierung besitzen, vertreten. Dieses vergleichsweise plötzliche Auftreten der „modernen“ Fauna wird als Kambrische Explosion bezeichnet.
Fossilien vielzelliger Tiere liegen also mit einiger Sicherheit nicht früher als im Ediacarium, nach dem Ende der Marinoischen Eiszeit, vor. Da aber die ersten Vertreter als weichhäutige Individuen sehr geringer Körpergröße kaum fossilierbar gewesen sein werden (und, selbst wenn sie tatsächlich entdeckt würden, wohl zu wenige Merkmale für eine abgesicherte Zuordnung besäßen), erscheint auch nach den Abschätzungen anhand der Methode der molekularen Uhr eine tatsächliche Entstehung bereits im Tonium, vor dem Eiszeitalter des Cryogeniums, durchaus nicht unwahrscheinlich.
Literatur
- Hynek Burda, Gero Hilken, Jan Zrzavý: Systematische Zoologie. UTB, Stuttgart 2008, ISBN 978-3-8252-3119-4.
- Peter Ax: Multicellular Animals: A new Approach to the Phylogenetic Order in Nature. Springer Verlag, 2012. ISBN 978 3642801143.
- Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere. 2. Auflage. Elsevier, Spektrum Akademischer Verlag, München 2007, ISBN 978-3-8274-1575-2.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 03.06. 2026